Representative Types
Representative Types
This section describes some representatives of each large protozoan phylum to provide a basis for comparing the groups and an idea of the diversity of protozoa. Forms such as Amoeba and Paramecium, although large and easy to obtain for study, are not wholly representative because their life histories are somewhat simpler than those of other members of their respective groups.
Phylum Sarcomastigophora
The Sarcomastigophora includes both protozoa that move by flagella (Mastigophora) and those that move by pseudopodia (Sarcodina). These characteristics are not mutually exclusive; some mastigophorans (flagellates) can form and use pseudopodia, and some sarcodines have flagellated stages in their life cycles.
Subphylum Mastigophora: The Flagellated Protozoa
Although some flagellates can form pseudopodia, their primary means of locomotion is by one or more flagella. The group is divided into phytoflagellates (class Phytomastigophorea), which usually have chlorophyll and are thus plantlike, and zooflagellates (class Zoomastigophorea), which do not have chlorophyll, are either holozoic or saprozoic, and thus are animal-like.
Phytoflagellates: Phytoflagellates usually have one or two (sometimes four) flagella and chloroplasts, which contain the pigments used in photosynthesis. They are mostly free living and include such familiar forms as Euglena, Chlamydomonas, Peranema, Volvox, and the dinoflagellates. Peranema (Figure 11-11) is related to Euglena but is a colorless phytoflagellate with holozoic nutrition. Chilomonas is another common form that is an important food item for amebas. Some flagellates are colonial, living in groups of zooids (each individual in a colonial animal or protozoan is a zooid). In some species the number of zooids per colony is characteristic (Figure 11-11).
Among the most interesting of all phytoflagellates are dinoflagellates. They have a longitudinal and an equatorial flagellum, each borne at least partly in grooves on the body. The body may be naked or covered by cellulose plates or valves. Most dinoflagellates have brown or yellow chromatophores, although some are colorless. Many species, both colorless and pigmented, can ingest prey through a mouth region between the plates near the posterior area of the body.Ceratium (Figure 11-11), for example, has a thick covering with long spines, into which the body extends, but it can catch food with posterior pseudopodia and ingest it between the flexible plates in the posterior groove. Noctiluca (Figure 11-11), a colorless dinoflagellate, is a voracious predator and has a long, motile tentacle, near the base of which its single, short flagellum emerges. Noctiluca is one of many marine organisms that can produce light (bioluminescence). Several groups of phytoflagellates are planktonic primary producers in freshwater and marine environments; however, dinoflagellates are the most important, particularly in the sea. Zooxanthellae are dinoflagellates that live in mutualistic association in the tissues of certain invertebrates, including other protozoa, sea anemones, horny and stony corals, and clams. The association with stony corals is of ecological and economic importance because only corals with symbiotic zooxanthellae can form coral reefs (Radiate Animals).
Euglena viridis: Euglena viridis (Figure 11-12) is a flagellate commonly studied in introductory zoology courses. Its natural habitat is freshwater streams and ponds where there is considerable vegetation. The organisms are spindle shaped and about 60 µm long, but some species of Euglena are smaller and some larger (E. oxyuris is 500 µm long). Just beneath the outer membrane of Euglena are proteinaceous strips and microtubules that form a pellicle. In Euglena the pellicle is flexible enough to permit bending, but in other euglenids it may be more rigid. A flagellum extends from a flask-shaped reservoir at the anterior end, and another, short flagellum ends within the reservoir. A kinetosome is found at the base of each flagellum, and a contractile vacuole empties into the reservoir. A red eyespot, or stigma, apparently functions in orientation to light. Within the cytoplasm are oval chloroplasts that bear chlorophyll and give the organism its greenish color. Paramylon bodies of various shapes are masses of a starchlike food storage material.
Nutrition of Euglena is normally autotrophic (holophytic), but if kept in the dark the organism makes use of saprozoic nutrition, absorbing nutrients through its body surface. Mutants of Euglena can be produced that have permanently lost their photosynthetic ability. Although Euglena does not ingest solid food, some euglenids are phagotrophic. Peranema has a cytostome that opens alongside its flagellar reservoir.
Euglena reproduces by binary fission and can encyst to survive adverse environmental conditions.
Volvox globator Volvox (Figure 11- 13) is a multicellular phytoflagellate that contains separate somatic and reproductive cells. It is often studied in introductory courses because its mode of development is somewhat similar to embryonic development of some metazoa. The order to which Volvox belongs (Volvocida) includes many freshwater flagellates, mostly green, with a cellulose cell wall through which two short flagella project. Many are colonial forms (Figure 11-11, Pandorina, Eudorina, Gonium), in which a single organism contains more than one cell but separate somatic and reproductive types do not exist.
Volvox (Figure 11-13) is a green, hollow sphere that may reach a diameter of 0.5 to 1 mm. A single organism contains many thousands of zooids (up to 50,000) embedded in the gelatinous surface of a jelly ball. Each cell is much like a euglenid, with a nucleus, a pair of flagella, a large chloroplast, and a red stigma. Adjacent cells are connected with each other by cytoplasmic strands. At one pole (usually in front as the colony moves), the stigmata are a little larger. Coordinated action of the flagella causes the colony to move by rolling over and over.
In Volvox we have a division of labor to the extent that most of the zooids are somatic cells concerned with nutrition and locomotion, and a few germ cells located in the posterior half are responsible for reproduction. Reproduction is asexual or sexual. In either case only certain zooids located around the equator or in the posterior half take part.
Asexual reproduction in Volvox occurs by repeated mitotic division of one of the germ cells to form a hollow sphere of cells, with the flagellated ends of the cells inside. The sphere then turns itself inside out to form a daughter colony similar to the parent colony. Several daughter colonies are formed inside the parent colony before they escape by rupture of the parent.
In sexual reproduction some of the zooids differentiate into macrogametes or microgametes (Figure 11-13). Macrogametes are fewer and larger and are loaded with food for nourishment of the young organism. Microgametes, by repeated division, form bundles or balls of small flagellated sperm that leave the mother organism when they mature and swim about to find a mature ovum. After fertilization, the zygote secretes a hard, spiny, protective shell around itself. When released by the breaking up of a parent, a zygote remains quiescent during the winter. Within its shell the zygote undergoes repeated division, producing a small organism that breaks out in the spring. A number of asexual generations may follow, during the summer, before sexual reproduction occurs again.
Zooflagellates: Zooflagellates are all colorless, lack chromoplasts, and have holozoic or saprozoic nutrition. Most are symbiotic.
Some of the most important protozoan parasites are zooflagellates. Many of them belong to the genus Trypanosoma (Figure 11-14) and live in the blood of fish, amphibians, reptiles, birds, and mammals. Some are nonpathogenic, but others produce severe diseases in humans and domestic animals. Trypanosoma brucei gambiense and T. brucei rhodesiense cause African sleeping sickness in humans, and T. brucei brucei causes a related disease in domestic animals. Trypanosomes are transmitted by tsetse flies (Glossina spp.). Trypanosoma b. rhodesiense, the more virulent of the sleeping sickness trypanosomes, and T. b. brucei have natural reservoirs (antelope and other wild mammals) that are apparently not harmed by the parasites. Some 10,000 new cases of human sleeping sickness are diagnosed each year, of which about half are fatal, and many of the remainder sustain permanent brain damage.
Trypanosoma cruzi causes Chagas disease in humans in Central America and South America. It is transmitted by “kissing bugs” (Triatominae), a name arising from the bug’s habit of biting its sleeping victim on the face. Acute Chagas disease is most common and severe among children less than five years old, while the chronic disease is seen most often in adults. Symptoms are primarily a result of central and peripheral nervous dysfunction. Two to three million people in South and Central America show chronic Chagas disease, and 45,000 of these die each year.
Several species of Leishmania cause disease in humans. Infection with some species may result in a serious visceral disease affecting especially the liver and spleen; others can cause disfiguring lesions in the mucous membranes of the nose and throat, and the least serious result is a skin ulcer. Leishmania spp. are transmitted by sand flies. Visceral leishmaniasis and cutaneous leishmaniasis are common in parts of Africa and Asia, and the mucocutaneous form is found in Central America and South America.
Trichomonas spp. (Figure 11.14) are symbiotic. Pentatrichomonas hominis lives in the cecum and colon of humans and Trichomonas tenax lives the mouth; they apparently cause no disease. Trichomonas vaginalis inhabits the urogenital tract of humans, is transmitted venereally, and is a common culprit in vaginitis. Other species of Trichomonadida are widely distributed through all classes of vertebrates and many invertebrates.
Giardia lamblia often causes no disease in the intestine of humans but sometimes may produce severe diarrhea. It is transmitted through fecal contamination and is cosmopolitan in distribution.
Subphylum Sarcodina
Superclass Rhizopoda Amoeba proteus The most commonly studied species of ameba is Amoeba proteus. These amebas live in slow streams and ponds of clear water, often in shallow water on aquatic vegetation or on sides of ledges. They are rarely found free in water, for they require a substratum on which to crawl. They have an irregular shape because lobopodia may be formed at any point on their bodies. They are colorless and about 250 to 600 µm in greatest diameter. Unlike Euglena, the pellicle consists only of a cell membrane. Ectoplasm and endoplasm are prominent. Organelles such as nucleus, contractile vacuole, food vacuoles, and small vesicles can be observed easily with a light microscope. Amebas live on algae, protozoa, rotifers, and even other amebas, upon which they feed by phagocytosis. An ameba can live for many days without food but decreases in volume during starvation. The time necessary for the digestion by a food vacuole varies with the kind of food but is usually around 15 to 30 hours. When an ameba reaches full size, it divides by binary fission with typical mitosis.
Other Rhizopoda There are many species of amebas; for example, A. verrucosa has short pseudopodia; Chaos carolinense (Pelomyxa carolinensis) is several times as large as A. proteus; and A. radiosa has many slender pseudopodia.
There are many entozoic amebas, most of which live in the intestines of humans or other animals. Two common genera are Endamoeba and Entamoeba. Endamoeba blattae is an endocommensal in the intestine of cockroaches, and related species are found in termites. Entamoeba histolytica is the most important rhizopod parasite of humans. It lives in the large intestine and on occasion can invade the intestinal wall by secreting enzymes that attack the intestinal lining. If this occurs, a serious and sometimes fatal amebic dysentery may result. The organisms may be carried by the blood to the liver and other organs and cause abscesses there. Many infected persons show few or no symptoms but are carriers, passing cysts in their feces. Diagnosis is complicated by the existence of a nonpathogenic species, E. dispar, which is morphologically identical to E. histolytica. Infection is spread by contaminated water or food containing cysts. Entamoeba histolytica is found throughout the world, but clinical amebiasis is most prevalent in tropical and subtropical areas.
Other species of Entamoeba found in humans are E. coli in the intestine and E. gingivalis in the mouth. Neither of these species is known to cause disease.
Not all rhizopods are “naked” as are amebas. Some have their delicate plasma membrane covered with a protective test or shell. Arcella and Difflugia (Figure 11-15) are common sarcodines. They have a test of secreted siliceous or chitinoid material that may be reinforced with grains of sand. They move by means of pseudopodia that project from openings in the shell.
Foraminiferans (class Granuloreticulosea) are an ancient group of shelled rhizopods found in all oceans, with a few in fresh and brackish water. Most foraminiferans live on the ocean floor in incredible numbers, having perhaps the largest biomass of any animal group on earth. Their tests are of numerous types (Figure 11-15 and 11-16). Most tests are many chambered and are made of calcium carbonate, although they sometimes use silica, silt, and other foreign materials. Slender pseudopodia extend through openings in the test, then branch and run together to form a protoplasmic net (reticulopodia) in which they ensnare their prey. Here captured prey is digested, and digested products are carried into the interior by flowing protoplasm. Life cycles of foraminiferans are complex, for they have multiple fission and alternation of haploid and diploid generations (intermediary meiosis).
Some slime molds (class Eumycetozoa),
especially Dictyostelium
discoideum, have been studied intensively
because of their fascinating
developmental cycle. Under natural
conditions this species lives in forest
detritus throughout the world. It feeds
on bacteria and reproduces by binary
fission as long as the food supply is
plentiful. When food runs short, however,
the amebas are attracted to each
other, streaming toward a central point
to form a pseudoplasmodium (large
mass of discrete cells). Under the same
conditions, some species actually fuse
to become a large multinucleate individual
(plasmodium). The pseudoplasmodium
of Dictyostelium may
migrate some distance to a favorable
location, where it forms a stalk with a
fruiting body on top (Figure 11-17). It
forms resistant cysts within the fruiting
body, which are widely dispersed
upon rupture of the fruiting body.
Many details about development,
genetics, and biochemistry of these
organisms are known.
Superclass Actinopoda Actinopoda is composed of the mostly freshwater class Heliozoea and three marine classes: Acantharea, Phaeodarea, and Polycystinea. Members of the marine classes are commonly called radiolarians. All have axopodia, and, except for some heliozoeans, they have tests (Figure 11-18). These protozoa are beautiful little organisms.
Biological characteristics of freshwater Heliozoea are somewhat better known than those of other actinopods. Examples are Actinosphaerium, which is about 1 mm in diameter and can be seen with the unassisted eye, and Actinophrys (Figure 11-15), only 50 µm in diameter; neither has a test. Clathrulina (Figure 11-15) secretes a latticed test.
The oldest known protozoa are found among the radiolarians. Radiolarians are nearly all pelagic (live in open water). Most of them are planktonic in shallow water, although some live in deep water. Their highly specialized skeletons are intricate in form and of great beauty (Figure 11-18). The body is divided by a central capsule that separates inner and outer zones of cytoplasm. The central capsule, which may be spherical, ovoid, or branched, is perforated to allow cytoplasmic continuity. The skeleton is made of silica, strontium sulfate, or a combination of silica and organic matter and usually has a radial arrangement of spines that extend through the capsule from the center of the body. At the surface a shell may be fused with the spines. Around the capsule is a frothy mass of cytoplasm from which axopodia arise . These are sticky to catch prey, which are carried by the streaming protoplasm to the central capsule to be digested. The ectoplasm on one side of the axial rod moves outward, or toward the tip, while on the other side it moves inward, or toward the test.
Radiolarians may have one or many nuclei. Their life history is not completely known, but binary fission, budding, and sporulation have been observed in them.
Role of Sarcodina in Building Earth Deposits Foraminiferans and radiolarians have existed since Precambrian times and have left excellent fossil records. In many instances their hard shells have been preserved unaltered. Many extinct species closely resemble those of the present day. They were especially abundant during the Cretaceous and Tertiary periods. Some were among the largest protozoa that have ever existed, measuring up to 100 mm (about 4 in) or more in diameter.
For untold millions of years tests of dead foraminiferans have been sinking to the bottom of the ocean, building up a characteristic ooze rich in lime and silica. About one-third of the sea bottom is covered with ooze that is made up of shells of the genus Globigerina. This ooze is especially abundant in the Atlantic Ocean.
Radiolarians (Figure 11-18), with their less soluble siliceous shells, are usually found at greater depths (4600 to 6100 meters), mainly in the Pacific and Indian oceans. Radiolarian ooze probably covers about 5 to 8 million square kilometers to a thickness of 700 to 4000 m. Under certain conditions, radiolarian ooze forms rocks (chert). Many fossil radiolarians are found in Tertiary rocks of California.
great pyramids of Egypt were made from stone quarried from limestone beds that were formed by a very large foraminiferan population that flourished during the early Tertiary period.
Since fossil foraminiferans and radiolarians can be found in well drillings, their identification is often important to oil geologists for correlation of rock strata.
Phylum Apicomplexa
All apicomplexans are endoparasites, and their hosts are found in many animal phyla. The presence of a certain combination of organelles, the apical complex, distinguishes this phylum (Figure 11-19A). The apical complex is usually present only in certain developmental stages of the organisms; for example, merozoites and sporozoites (Figure 11-20). Some structures, especially the rhoptries and micronemes, apparently aid in penetrating the host’s cells or tissues.
Locomotor organelles are less obvious in this group than in other protozoa. Pseudopodia occur in some intracellular stages, and gametes of some species are flagellated. Tiny contractile fibrils can form waves of contraction across the body surfaces to propel the organism through a liquid medium.
The life cycle usually includes both asexual and sexual reproduction, and sometimes an invertebrate intermediate host. At some point in the life cycle, the organisms develop a spore (oocyst), which is infective for the next host and is often protected by a resistant coat.
Class Sporozoa
The most important class of phylum Apicomplexa, Sporozoa, contains three subclasses: Gregarinia, Coccidia, and Piroplasmia. Gregarines are common parasites of invertebrates, but they are of little economic significance. Piroplasms are of some veterinary importance; for example, Babesia bigemina causes Texas red-water fever in cattle. Humans are occasionally infected with species of Babesia normally parasitic in other animals.
Subclass Coccidia Coccidia are intracellular parasites in invertebrates and vertebrates, and the group includes species of very great medical and veterinary importance.
Eimeria. The name “coccidiosis” is generally applied only to infections with Eimeria or Isospora. Humans can be infected with species of Isospora, but there is usually little disease. However, Isospora infections can be very serious in AIDS patients. Some species of Eimeria may cause serious disease in some domestic animals. Symptoms usually include severe diarrhea or dysentery.
Eimeria tenella is often fatal to young fowl, producing severe pathogenesis in the intestine. The organisms undergo schizogony in the intestinal cells, finally producing gametes. After fertilization the zygote forms an oocyst that passes out of its host in the feces (Figure 11-19B). Sporogony occurs within the oocyst outside the host, producing eight sporozoites in each oocyst. Infection occurs when a new host accidentally ingests a sporulated oocyst and the sporozoites are released by digestive enzymes.
Toxoplasma gondii. A similar life cycle occurs in Toxoplasma gondii, a parasite of cats, but this species produces extraintestinal stages as well. When rodents, cattle, sheep, humans, many other mammals, or even birds, ingest sporozoites, the sporozoites cross from the intestine and begin rapid, asexual reproduction in a variety of tissues. As the host mounts an immune response (see Immunity), reproduction of the zoites slows, and they become enclosed in tough tissue cysts. The zoites, now called bradyzoites, accumulate in large numbers in each tissue cyst. Bradyzoites are infective for other hosts, including cats, where they can initiate the intestinal cycle in a cat that eats infected prey. Bradyzoites can remain viable and infective for months or years, and it is estimated that onethird of the world’s human population carries tissue cysts containing bradyzoites in their body. Up to 50% of the human population of the United States are infected. The normal route of infection for humans is apparently consumption of infected meat that is insufficiently cooked.
In humans Toxoplasma causes little or no ill effects except in AIDS patients or in women infected during pregnancy, particularly in the first trimester. Such infection greatly increases the chances of a birth defect in the baby; it is now believed that 2% of all mental retardation in the United States is a result of congenital toxoplasmosis. Toxoplasmosis can also be a serious disease in persons who are immunosuppressed, either with drugs or by AIDS. In such patients rupture of a tissue cyst, which would be contained easily in a person with a normal immune system, becomes a source of life-threatening infection.
Plasmodium: The Malarial Organism: The best known of the coccidians is Plasmodium spp., causative organisms of the most important infectious disease of humans: malaria. Malaria is a very serious disease, difficult to control and widespread, particularly in tropical and subtropical countries. Four species of Plasmodium infect humans. Although each species produces its own peculiar clinical picture, all four have similar cycles of development in their hosts (Figure 11-20).
The parasite is carried by mosquitoes (Anopheles), and sporozoites are injected into a human with the insect’s saliva during its bite. Sporozoites penetrate liver cells and initiate schizogony. The products of this division then enter other liver cells to repeat the schizogonous cycle, or in P. falciparum they penetrate the red blood cells after only one cycle in the liver. The period when the parasites are in the liver is the incubation period, and it lasts from 6 to 15 days, depending on the species of Plasmodium.
Merozoites released as a result of liver schizogony enter red blood cells, where they begin a series of schizogonous cycles. When they enter red blood cells, they become ameboid trophozoites, feeding on hemoglobin. The end product of the parasite’s digestion of hemoglobin is a dark, insoluble pigment: hemozoin. Hemozoin accumulates in the host cell, is released when the next generation of merozoites is produced, and eventually accumulates in the liver, spleen, or other organs. The trophozoite within a red blood cell grows and undergoes schizogony, producing 6 to 36 merozoites, which, depending on the species, burst forth to infect new red cells. When a red blood cell containing merozoites bursts, it releases the parasite’s metabolic products, which have accumulated there. Release of these foreign substances into the patient’s circulation results in the chills and fever characteristic of malaria.
Since the populations of schizonts maturing in red blood cells are synchronized to some degree, the episodes of chills and fever have a periodicity characteristic of the particular species of Plasmodium. In P. vivax (benign tertian) malaria and P. ovale malaria, episodes occur every 48 hours; in P. malariae (quartan) malaria, every 72 hours; and in P. falciparum (malignant tertian) malaria, about every 48 hours, although synchrony is less well defined in this species. People usually recover from infections with the first three species, but mortality may be high in untreated cases of P. falciparum infection. Sometimes grave complications, such as cerebral malaria, occur. Unfortunately, P. falciparum is the most common species, accounting for 50% of all malaria in the world. Certain genes, for example the gene for sickle cell hemoglobin, confer some resistance to malaria on people that carry them.
After some cycles of schizogony in red blood cells, infection of new cells by some of the merozoites results in production of microgametocytes and macrogametocytes rather than another generation of merozoites. When gametocytes are ingested by a mosquito feeding on a patient’s blood, they mature into gametes, and fertilization occurs. The zygote becomes a motile ookinete, which penetrates the stomach wall of the mosquito and becomes an oocyst. Within the oocyst, sporogony occurs, and thousands of sporozoites are produced. The oocyst ruptures, and the sporozoites migrate to the salivary glands, from which they are transferred to a human by a bite of the mosquito. Development in a mosquito requires 7 to 18 days but may be longer in cool weather.
Elimination of mosquitoes and their breeding places by insecticides, drainage, and other methods has been effective in controlling malaria in some areas. However, difficulties in carrying out such activities in remote areas and areas suffering civil unrest, and acquisition of resistance to insecticides by mosquitoes and to antimalarial drugs by Plasmodium (especially P. falciparum), mean that malaria will be a serious disease of humans for a long time to come.
Other species of Plasmodium parasitize birds, reptiles, and mammals. Those of birds are transmitted chiefly by Culex mosquitoes.
Phylum Ciliophora
Ciliates are a large and interesting group, with a great variety of forms living in all types of freshwater and marine habitats. They are the most structurally complex and diversely specialized of all protozoan groups. The majority are free living, but some are commensal or parasitic. They are usually solitary and motile, but some are sessile and some colonial. There is great diversity of shape and size. In general, they are larger than most other protozoa, but they range from very small (10 to 12 µm) up to 3 mm long. All have cilia that beat in a coordinated rhythmical manner, although the arrangement of the cilia may vary and some lack cilia as adults.
Ciliates are always multinucleate, possessing at least one macronucleus and one micronucleus, but varying from one to many of either type. The macronuclei are apparently responsible for metabolic and developmental functions and for maintaining all the visible traits, such as the pellicular apparatus. Macronuclei vary in shape among the different species (Figures 11-21 and 11-23). Micronuclei participate in sexual reproduction and give rise to macronuclei after exchange of micronuclear material between individuals. Micronuclei divide mitotically, and macronuclei divide amitotically.
The pellicle of ciliates may consist only of a cell membrane or in some species may form a thickened armor. Cilia are short and usually arranged in longitudinal or diagonal rows. Cilia may cover the surface of the organism or may be restricted to the oral region or to certain bands. In some forms cilia are fused into a sheet called an undulating membrane or into smaller membranelles, both used to propel food into the cytopharynx (gullet). In other forms there may be fused cilia forming stiffened tufts called cirri, often used in locomotion by the creeping ciliates (Figure 11-21).
An apparently structural system of fibers, in addition to the kinetosomes, makes up the infraciliature, just beneath the pellicle (Figure 11-22). Each cilium terminates beneath the pellicle in its kinetosome, and from each kinetosome a fibril arises and passes along beneath the row of cilia, joining with the other fibrils of that row. The cilia, kinetosomes, and other fibrils of that ciliary row make up what is called a kinety (Figure 11-22). All ciliates seem to have kinety systems, even those that lack cilia at some stage. The infraciliature apparently does not coordinate ciliary beat, as formerly thought. Coordination of ciliary movement seems to be by waves of depolarization of the cell membrane moving down the organism, similar to the phenomenon in a nerve impulse.
Most ciliates are holozoic. Most of them possess a cytostome (mouth) that in some forms is a simple opening and in others is connected to a gullet or ciliated groove. The mouth in some is strengthened with stiff, rodlike trichites for swallowing larger prey; in others, such as paramecia, ciliary water currents carry microscopic food particles toward the mouth. Didinium has a proboscis for engulfing paramecia on which it feeds (Figure 11-9). Suctorians paralyze their prey and then ingest the contents through tubelike tentacles by a complex feeding mechanism that apparently combines phagocytosis with a sliding filament action of microtubules in the tentacles (Figure 11-9).
Some ciliates have curious small bodies in their ectoplasm between the bases of the cilia. Examples are trichocysts (Figure 11-22 and 11-23) and toxicysts. Upon mechanical or chemical stimulation, these bodies explosively expel a long, threadlike structure. The mechanism of expulsion is unknown. The function of trichocysts is thought to be defensive. When attacked by a Didinium, a paramecium expels its trichocysts but to no avail. Toxicysts, however, release a poison that paralyzes the prey of carnivorous ciliates. Toxicysts are structurally quite distinct from trichocysts. Many dinoflagellates have structures very similar to trichocysts.
Among the more striking and familiar of ciliates are Stentor (Gr. herald with a loud voice), trumpet shaped and solitary, with a beadshaped macronucleus (Figure 11-21); Vorticella (L. dim. of vortex, a whirlpool), bell shaped and attached by a contractile stalk (Figure 11-21); and Euplotes (Gr. eu, true, good, + ploter, swimmer) with a flattened body and groups of fused cilia (cirri) that function as legs.
Paramecium: A Representative Ciliate
Paramecia are usually abundant in ponds or sluggish streams containing aquatic plants and decaying organic matter.
Form and Function Paramecia are often described as slipper shaped. Paramecium caudatum is 150 to 300 µm in length and is blunt anteriorly and somewhat pointed posteriorly (Figure 11-23). The organism has an asymmetrical appearance because of the oral groove, a depression that runs obliquely backward on the ventral side.
The pellicle is a clear, elastic membrane that may be ornamented by ridges or papillalike projections (Figure 11-22), and its entire surface is covered with cilia arranged in lengthwise rows. Just below the pellicle is the thin clear ectoplasm that surrounds the larger mass of granular endoplasm (Figure 11-23). Embedded in ectoplasm just below the surface are spindleshaped trichocysts, which alternate with the bases of cilia. The infraciliature can be seen only with special fixing and staining methods.
The cytostome at the end of the oral groove leads into a tubular cytopharynx, or gullet. Along the gullet an undulating membrane of modified cilia keeps food moving. Fecal material is discharged through a cytoproct posterior to the oral groove (Figure 11-23). Within the endoplasm are food vacuoles containing food in various stages of digestion. There are two contractile vacuoles, each consisting of a central space surrounded by several radiating canals (Figure 11-23) that collect fluid and empty it into the central vacuole.
Paramecium caudatum has two nuclei: a large kidney-shaped macronucleus and a smaller micronucleus fitted into the depression of the former. These can usually be seen only in stained specimens. The number of micronuclei varies in different species; for example, P. multimicronucleatum may have as many as seven.
Paramecia are holozoic, living on bacteria, algae, and other small organisms. Cilia in the oral groove sweep food particles in the water into the cytostome, from which point they are carried into the cytopharynx by the undulating membrane. From the cytopharynx food is collected into a food vacuole that is constricted into the endoplasm. Food vacuoles circulate in a definite course through the cytoplasm while the food is being digested by enzymes from the endoplasm. The indigestible part of the food is ejected through the cytoproct.
The body is elastic, allowing it to bend and squeeze its way through narrow places. Its cilia can beat either forward or backward, so that the organism can swim in either direction. The cilia beat obliquely, causing the organism to rotate on its long axis. In the oral groove the cilia are longer and beat more vigorously than the others so that the anterior end swerves aborally. As a result of these factors, the organism moves forward in a spiral path (Figure 11-24A).
When a ciliate, such as a paramecium, comes in contact with a barrier or a disturbing chemical stimulus, it reverses its cilia, backs up a short distance, and swerves the anterior end as it pivots on its posterior end. This behavior is called an avoiding reaction (Figure 11-24B). A paramecium may continue to change its direction to keep itself away from the noxious stimulus, and it may react in a similar fashion to keep itself within the zone of an attractant. A paramecium may also change its swimming speed. How does a paramecium “know” when to change directions or swimming speed? Interestingly, the reactions of the organism depend on the effects of the stimulus on the electrical potential difference across its cell membrane. Paramecia slightly hyperpolarize in attractants and depolarize in repellents that produce the avoiding reaction. Hyperpolarization increases the rate of the forward ciliary beat, and depolarization results in ciliary reversal and backward swimming.
Reproduction Paramecia reproduce only by binary fission across kineties (ciliary rows) but have certain forms of sexual phenomena called conjugation and autogamy.
In binary fission the micronucleus divides mitotically into two daughter micronuclei, which move to opposite ends of the cell (Figure 11-25). The macronucleus elongates and divides amitotically.
Conjugation occurs at intervals in
ciliates. Conjugation is the temporary
union of two individuals to exchange
chromosomal material (Figure 11-26).
During the union the macronucleus disintegrates
and the micronucleus of each
individual undergoes meiosis, giving
rise to four haploid micronuclei, three of
which degenerate (Figure 11-26A to D).
The remaining micronucleus then divides
into two haploid pronuclei, one of
which is exchanged with the other conjugant.
The pronuclei fuse to restore the
diploid number of chromosomes, followed
by several more nuclear events
detailed in Figure 11-26. Following this
complicated process, the organisms may
continue to reproduce by binary fission
without the necessity of conjugation.
The result of conjugation is similar to that of zygote formation, for each exconjugant contains hereditary material from two individuals. The advantage of sexual reproduction is that it permits gene recombinations, thus increasing genetic variation in the population. Although ciliates in clone cultures can apparently reproduce repeatedly and indefinitely without conjugation, the stock seems eventually to lose vigor. Conjugation restores vitality to a stock. Seasonal changes or a deteriorating environment will usually stimulate sexual reproduction.
Autogamy is a process of selffertilization that is similar to conjugation except that there is no exchange of nuclei. After the disintegration of the macronucleus and the meiotic divisions of the micronucleus, two haploid pronuclei fuse to form a synkaryon that is completely homozygous (Principles of Genetics:A Review).
Symbiotic Ciliates
Many symbiotic ciliates live as commensals, but some can be harmful to their hosts. Balantidium coli lives in the large intestine of humans, pigs, rats, and many other mammals (Figure 11-27). There seem to be hostspecific strains, and the organism is not easily transmitted from one species to another. Transmission is by fecal contamination of food or water. Usually the organisms are not pathogenic, but in humans they sometimes invade the intestinal lining and cause a dysentery similar to that caused by Entamoeba histolytica. The disease can be serious and even fatal. Infections are common in parts of Europe, Asia, and Africa but are rare in the United States.
Other species of ciliates live in other hosts. Entodinium (Figure 11-27) belongs to a group that has very complex structure and lives in the digestive tract of ruminants, where they may be very abundant. Nyctotherus live in the colon of frogs and toads. In aquarium and wild freshwater fishes, Ichthyophthirius causes a disease known to many fish culturists as “ick.” Untreated, it can cause much loss of exotic fishes.
Suctorians
Suctorians are ciliates in which the young possess cilia and are free swimming, and the adults grow a stalk for attachment, become sessile, and lose their cilia. They have no cytostome but feed by long, slender, tubelike tentacles. The suctorian captures living prey, usually a ciliate, by the tip of one or more tentacles and paralyzes it. The cytoplasm of the prey then flows through the attached tentacles, forming food vacuoles in the feeding suctorian (Figure 11-9).
One of the best places to find freshwater suctorians is in algae that grow on the carapace of turtles. Common genera of suctorians found there are Anarma (without stalk or test) and Squalorophrya (with stalk and test). Other freshwater representatives are Podophrya (Figure 11-9) and Dendrosoma. Acinetopsis and Ephelota are saltwater forms. Suctorian parasites include Trichophrya, the species of which are found on a variety of invertebrates and freshwater fish; Allantosoma, which live in the intestine of certain mammals; and Sphaerophrya, which are found in Stentor.
This section describes some representatives of each large protozoan phylum to provide a basis for comparing the groups and an idea of the diversity of protozoa. Forms such as Amoeba and Paramecium, although large and easy to obtain for study, are not wholly representative because their life histories are somewhat simpler than those of other members of their respective groups.
Phylum Sarcomastigophora
The Sarcomastigophora includes both protozoa that move by flagella (Mastigophora) and those that move by pseudopodia (Sarcodina). These characteristics are not mutually exclusive; some mastigophorans (flagellates) can form and use pseudopodia, and some sarcodines have flagellated stages in their life cycles.
Subphylum Mastigophora: The Flagellated Protozoa
Although some flagellates can form pseudopodia, their primary means of locomotion is by one or more flagella. The group is divided into phytoflagellates (class Phytomastigophorea), which usually have chlorophyll and are thus plantlike, and zooflagellates (class Zoomastigophorea), which do not have chlorophyll, are either holozoic or saprozoic, and thus are animal-like.
Phytoflagellates: Phytoflagellates usually have one or two (sometimes four) flagella and chloroplasts, which contain the pigments used in photosynthesis. They are mostly free living and include such familiar forms as Euglena, Chlamydomonas, Peranema, Volvox, and the dinoflagellates. Peranema (Figure 11-11) is related to Euglena but is a colorless phytoflagellate with holozoic nutrition. Chilomonas is another common form that is an important food item for amebas. Some flagellates are colonial, living in groups of zooids (each individual in a colonial animal or protozoan is a zooid). In some species the number of zooids per colony is characteristic (Figure 11-11).
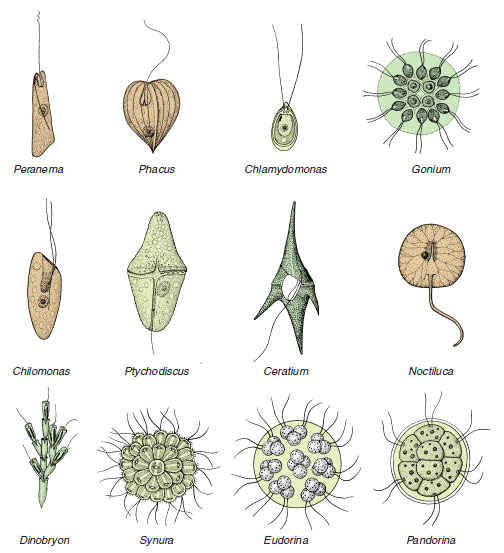 |
| Figure 11-11 Diversity among Phytomastigophorea. Pandorina, Eudorina, Synura, Gonium, and Dinobryon are colonial. Ptychodiscus, Ceratium, and Chlamydomas are dinoflagellates. Noctiluca, Peranema, and Chilomonas have no pigments and are not photosynthetic. Phacus has two flagella, one of which is very short, as in Euglena. |
Among the most interesting of all phytoflagellates are dinoflagellates. They have a longitudinal and an equatorial flagellum, each borne at least partly in grooves on the body. The body may be naked or covered by cellulose plates or valves. Most dinoflagellates have brown or yellow chromatophores, although some are colorless. Many species, both colorless and pigmented, can ingest prey through a mouth region between the plates near the posterior area of the body.Ceratium (Figure 11-11), for example, has a thick covering with long spines, into which the body extends, but it can catch food with posterior pseudopodia and ingest it between the flexible plates in the posterior groove. Noctiluca (Figure 11-11), a colorless dinoflagellate, is a voracious predator and has a long, motile tentacle, near the base of which its single, short flagellum emerges. Noctiluca is one of many marine organisms that can produce light (bioluminescence). Several groups of phytoflagellates are planktonic primary producers in freshwater and marine environments; however, dinoflagellates are the most important, particularly in the sea. Zooxanthellae are dinoflagellates that live in mutualistic association in the tissues of certain invertebrates, including other protozoa, sea anemones, horny and stony corals, and clams. The association with stony corals is of ecological and economic importance because only corals with symbiotic zooxanthellae can form coral reefs (Radiate Animals).
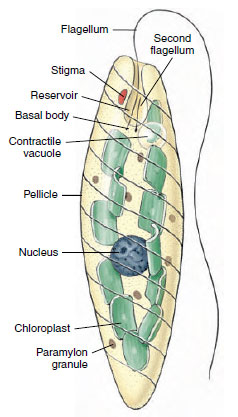 |
| Figure 11-12 Euglena. Features shown are a combination of those visible in living and stained preparations. |
Euglena viridis: Euglena viridis (Figure 11-12) is a flagellate commonly studied in introductory zoology courses. Its natural habitat is freshwater streams and ponds where there is considerable vegetation. The organisms are spindle shaped and about 60 µm long, but some species of Euglena are smaller and some larger (E. oxyuris is 500 µm long). Just beneath the outer membrane of Euglena are proteinaceous strips and microtubules that form a pellicle. In Euglena the pellicle is flexible enough to permit bending, but in other euglenids it may be more rigid. A flagellum extends from a flask-shaped reservoir at the anterior end, and another, short flagellum ends within the reservoir. A kinetosome is found at the base of each flagellum, and a contractile vacuole empties into the reservoir. A red eyespot, or stigma, apparently functions in orientation to light. Within the cytoplasm are oval chloroplasts that bear chlorophyll and give the organism its greenish color. Paramylon bodies of various shapes are masses of a starchlike food storage material.
Nutrition of Euglena is normally autotrophic (holophytic), but if kept in the dark the organism makes use of saprozoic nutrition, absorbing nutrients through its body surface. Mutants of Euglena can be produced that have permanently lost their photosynthetic ability. Although Euglena does not ingest solid food, some euglenids are phagotrophic. Peranema has a cytostome that opens alongside its flagellar reservoir.
Euglena reproduces by binary fission and can encyst to survive adverse environmental conditions.
Volvox globator Volvox (Figure 11- 13) is a multicellular phytoflagellate that contains separate somatic and reproductive cells. It is often studied in introductory courses because its mode of development is somewhat similar to embryonic development of some metazoa. The order to which Volvox belongs (Volvocida) includes many freshwater flagellates, mostly green, with a cellulose cell wall through which two short flagella project. Many are colonial forms (Figure 11-11, Pandorina, Eudorina, Gonium), in which a single organism contains more than one cell but separate somatic and reproductive types do not exist.
Volvox (Figure 11-13) is a green, hollow sphere that may reach a diameter of 0.5 to 1 mm. A single organism contains many thousands of zooids (up to 50,000) embedded in the gelatinous surface of a jelly ball. Each cell is much like a euglenid, with a nucleus, a pair of flagella, a large chloroplast, and a red stigma. Adjacent cells are connected with each other by cytoplasmic strands. At one pole (usually in front as the colony moves), the stigmata are a little larger. Coordinated action of the flagella causes the colony to move by rolling over and over.
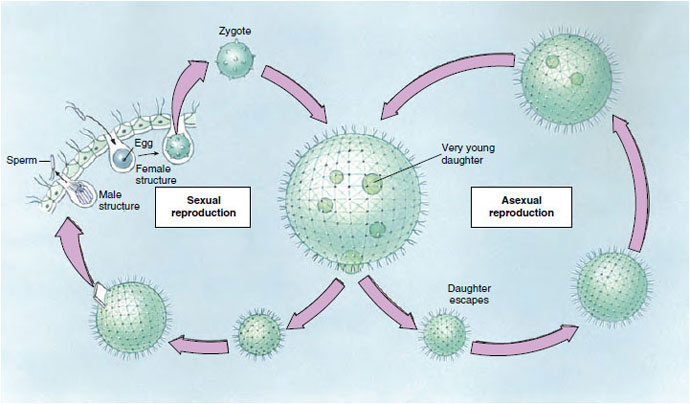 |
| Figure 11-13 Life cycle of Volvox. Asexual reproduction occurs in spring and summer when specialized diploid reproductive cells divide to form young organisms that remain in the mother organism until large enough to escape. Sexual reproduction occurs largely in autumn when haploid sex cells develop. The fertilized ova may encyst and so survive the winter, developing into a mature asexual organism in the spring. In some species the organisms have separate sexes; in others both eggs and sperm are produced in the same organism. |
In Volvox we have a division of labor to the extent that most of the zooids are somatic cells concerned with nutrition and locomotion, and a few germ cells located in the posterior half are responsible for reproduction. Reproduction is asexual or sexual. In either case only certain zooids located around the equator or in the posterior half take part.
Asexual reproduction in Volvox occurs by repeated mitotic division of one of the germ cells to form a hollow sphere of cells, with the flagellated ends of the cells inside. The sphere then turns itself inside out to form a daughter colony similar to the parent colony. Several daughter colonies are formed inside the parent colony before they escape by rupture of the parent.
In sexual reproduction some of the zooids differentiate into macrogametes or microgametes (Figure 11-13). Macrogametes are fewer and larger and are loaded with food for nourishment of the young organism. Microgametes, by repeated division, form bundles or balls of small flagellated sperm that leave the mother organism when they mature and swim about to find a mature ovum. After fertilization, the zygote secretes a hard, spiny, protective shell around itself. When released by the breaking up of a parent, a zygote remains quiescent during the winter. Within its shell the zygote undergoes repeated division, producing a small organism that breaks out in the spring. A number of asexual generations may follow, during the summer, before sexual reproduction occurs again.
Zooflagellates: Zooflagellates are all colorless, lack chromoplasts, and have holozoic or saprozoic nutrition. Most are symbiotic.
Some of the most important protozoan parasites are zooflagellates. Many of them belong to the genus Trypanosoma (Figure 11-14) and live in the blood of fish, amphibians, reptiles, birds, and mammals. Some are nonpathogenic, but others produce severe diseases in humans and domestic animals. Trypanosoma brucei gambiense and T. brucei rhodesiense cause African sleeping sickness in humans, and T. brucei brucei causes a related disease in domestic animals. Trypanosomes are transmitted by tsetse flies (Glossina spp.). Trypanosoma b. rhodesiense, the more virulent of the sleeping sickness trypanosomes, and T. b. brucei have natural reservoirs (antelope and other wild mammals) that are apparently not harmed by the parasites. Some 10,000 new cases of human sleeping sickness are diagnosed each year, of which about half are fatal, and many of the remainder sustain permanent brain damage.
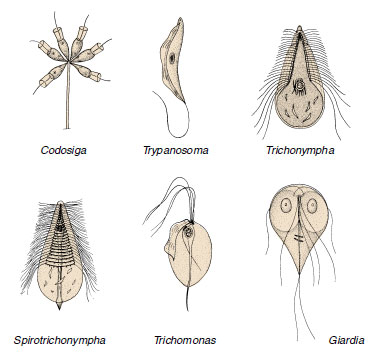 |
| Figure 11-14 Some Zoomastigophorea. Codosiga is a colonial flagellate with cells similar to those found in sponges (phylum Porifera). The others are all symbiotic. Trichonympha, Spirotrichonympha, and Trichomonas are commonly found in the gut of termites and wood roaches, where they help digest cellulose from the wood eaten by the insects. Species of Trichomonas are also found in humans. Trypanosoma is a parasite of various animals, and some species cause serious disease in humans and domestic animals. Giardia is an intestinal parasite of mammals that causes diarrhea in humans. |
Trypanosoma cruzi causes Chagas disease in humans in Central America and South America. It is transmitted by “kissing bugs” (Triatominae), a name arising from the bug’s habit of biting its sleeping victim on the face. Acute Chagas disease is most common and severe among children less than five years old, while the chronic disease is seen most often in adults. Symptoms are primarily a result of central and peripheral nervous dysfunction. Two to three million people in South and Central America show chronic Chagas disease, and 45,000 of these die each year.
Several species of Leishmania cause disease in humans. Infection with some species may result in a serious visceral disease affecting especially the liver and spleen; others can cause disfiguring lesions in the mucous membranes of the nose and throat, and the least serious result is a skin ulcer. Leishmania spp. are transmitted by sand flies. Visceral leishmaniasis and cutaneous leishmaniasis are common in parts of Africa and Asia, and the mucocutaneous form is found in Central America and South America.
Trichomonas spp. (Figure 11.14) are symbiotic. Pentatrichomonas hominis lives in the cecum and colon of humans and Trichomonas tenax lives the mouth; they apparently cause no disease. Trichomonas vaginalis inhabits the urogenital tract of humans, is transmitted venereally, and is a common culprit in vaginitis. Other species of Trichomonadida are widely distributed through all classes of vertebrates and many invertebrates.
Giardia lamblia often causes no disease in the intestine of humans but sometimes may produce severe diarrhea. It is transmitted through fecal contamination and is cosmopolitan in distribution.
Subphylum Sarcodina
Superclass Rhizopoda Amoeba proteus The most commonly studied species of ameba is Amoeba proteus. These amebas live in slow streams and ponds of clear water, often in shallow water on aquatic vegetation or on sides of ledges. They are rarely found free in water, for they require a substratum on which to crawl. They have an irregular shape because lobopodia may be formed at any point on their bodies. They are colorless and about 250 to 600 µm in greatest diameter. Unlike Euglena, the pellicle consists only of a cell membrane. Ectoplasm and endoplasm are prominent. Organelles such as nucleus, contractile vacuole, food vacuoles, and small vesicles can be observed easily with a light microscope. Amebas live on algae, protozoa, rotifers, and even other amebas, upon which they feed by phagocytosis. An ameba can live for many days without food but decreases in volume during starvation. The time necessary for the digestion by a food vacuole varies with the kind of food but is usually around 15 to 30 hours. When an ameba reaches full size, it divides by binary fission with typical mitosis.
Other Rhizopoda There are many species of amebas; for example, A. verrucosa has short pseudopodia; Chaos carolinense (Pelomyxa carolinensis) is several times as large as A. proteus; and A. radiosa has many slender pseudopodia.
There are many entozoic amebas, most of which live in the intestines of humans or other animals. Two common genera are Endamoeba and Entamoeba. Endamoeba blattae is an endocommensal in the intestine of cockroaches, and related species are found in termites. Entamoeba histolytica is the most important rhizopod parasite of humans. It lives in the large intestine and on occasion can invade the intestinal wall by secreting enzymes that attack the intestinal lining. If this occurs, a serious and sometimes fatal amebic dysentery may result. The organisms may be carried by the blood to the liver and other organs and cause abscesses there. Many infected persons show few or no symptoms but are carriers, passing cysts in their feces. Diagnosis is complicated by the existence of a nonpathogenic species, E. dispar, which is morphologically identical to E. histolytica. Infection is spread by contaminated water or food containing cysts. Entamoeba histolytica is found throughout the world, but clinical amebiasis is most prevalent in tropical and subtropical areas.
Other species of Entamoeba found in humans are E. coli in the intestine and E. gingivalis in the mouth. Neither of these species is known to cause disease.
Not all rhizopods are “naked” as are amebas. Some have their delicate plasma membrane covered with a protective test or shell. Arcella and Difflugia (Figure 11-15) are common sarcodines. They have a test of secreted siliceous or chitinoid material that may be reinforced with grains of sand. They move by means of pseudopodia that project from openings in the shell.
Foraminiferans (class Granuloreticulosea) are an ancient group of shelled rhizopods found in all oceans, with a few in fresh and brackish water. Most foraminiferans live on the ocean floor in incredible numbers, having perhaps the largest biomass of any animal group on earth. Their tests are of numerous types (Figure 11-15 and 11-16). Most tests are many chambered and are made of calcium carbonate, although they sometimes use silica, silt, and other foreign materials. Slender pseudopodia extend through openings in the test, then branch and run together to form a protoplasmic net (reticulopodia) in which they ensnare their prey. Here captured prey is digested, and digested products are carried into the interior by flowing protoplasm. Life cycles of foraminiferans are complex, for they have multiple fission and alternation of haploid and diploid generations (intermediary meiosis).
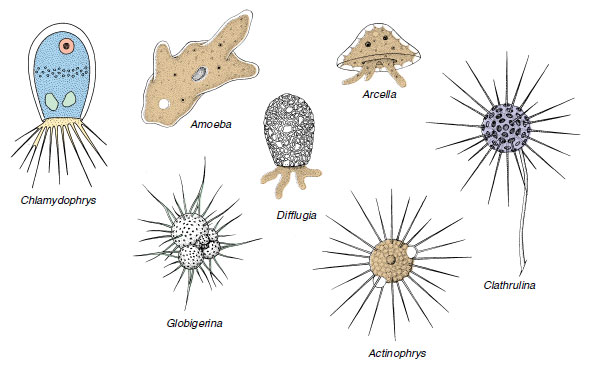 |
| Figure 11-15 Diversity among the Sarcodina. Difflugia, Arcella, and Amoeba belong to the rhizopod class Lobosea and have lobopodia. Chlamydophrys is in the class Filosea and has filopodia. The foraminiferan Globigerina belongs to the class Granuloreticulosea and shows reticulopodia. Actinophrys and Clathrulina are actinopod heliozoeans. They have axopodia. |
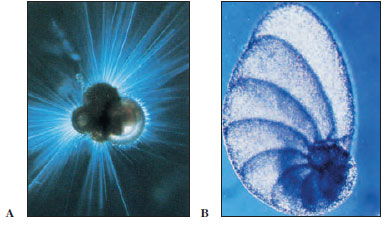 |
| Figure 11-16 A, Living foraminiferan, showing thin pseudopodia extending from test. B, Test of foraminiferan, Vertebralima striata. Foraminiferans (class Granuloreticulosea) are ameboid marine protozoa that secrete a calcareous, many- chambered test in which to live and then extrude protoplasm through pores to form a layer over the outside. The animal begins with one chamber, and as it grows, it secretes a succession of new and larger chambers, continuing this process throughout life. Many foraminiferans are planktonic, and when they die, their shells are added to the ooze on the ocean’s bottom. |
 |
| Figure 11-17 Fruiting bodies of three genera of plasmodial slime molds. A, Arcyria. B, Fuligo. C, Tubifera. |
Superclass Actinopoda Actinopoda is composed of the mostly freshwater class Heliozoea and three marine classes: Acantharea, Phaeodarea, and Polycystinea. Members of the marine classes are commonly called radiolarians. All have axopodia, and, except for some heliozoeans, they have tests (Figure 11-18). These protozoa are beautiful little organisms.
Biological characteristics of freshwater Heliozoea are somewhat better known than those of other actinopods. Examples are Actinosphaerium, which is about 1 mm in diameter and can be seen with the unassisted eye, and Actinophrys (Figure 11-15), only 50 µm in diameter; neither has a test. Clathrulina (Figure 11-15) secretes a latticed test.
 |
| Figure 11-18 Types of radiolarian tests (class Polycystinea). In his study of these beautiful forms collected on the famous Challenger expedition of 1872 to 1876, Haeckel proposed our present concepts of symmetry. |
The oldest known protozoa are found among the radiolarians. Radiolarians are nearly all pelagic (live in open water). Most of them are planktonic in shallow water, although some live in deep water. Their highly specialized skeletons are intricate in form and of great beauty (Figure 11-18). The body is divided by a central capsule that separates inner and outer zones of cytoplasm. The central capsule, which may be spherical, ovoid, or branched, is perforated to allow cytoplasmic continuity. The skeleton is made of silica, strontium sulfate, or a combination of silica and organic matter and usually has a radial arrangement of spines that extend through the capsule from the center of the body. At the surface a shell may be fused with the spines. Around the capsule is a frothy mass of cytoplasm from which axopodia arise . These are sticky to catch prey, which are carried by the streaming protoplasm to the central capsule to be digested. The ectoplasm on one side of the axial rod moves outward, or toward the tip, while on the other side it moves inward, or toward the test.
Radiolarians may have one or many nuclei. Their life history is not completely known, but binary fission, budding, and sporulation have been observed in them.
Role of Sarcodina in Building Earth Deposits Foraminiferans and radiolarians have existed since Precambrian times and have left excellent fossil records. In many instances their hard shells have been preserved unaltered. Many extinct species closely resemble those of the present day. They were especially abundant during the Cretaceous and Tertiary periods. Some were among the largest protozoa that have ever existed, measuring up to 100 mm (about 4 in) or more in diameter.
For untold millions of years tests of dead foraminiferans have been sinking to the bottom of the ocean, building up a characteristic ooze rich in lime and silica. About one-third of the sea bottom is covered with ooze that is made up of shells of the genus Globigerina. This ooze is especially abundant in the Atlantic Ocean.
Radiolarians (Figure 11-18), with their less soluble siliceous shells, are usually found at greater depths (4600 to 6100 meters), mainly in the Pacific and Indian oceans. Radiolarian ooze probably covers about 5 to 8 million square kilometers to a thickness of 700 to 4000 m. Under certain conditions, radiolarian ooze forms rocks (chert). Many fossil radiolarians are found in Tertiary rocks of California.
great pyramids of Egypt were made from stone quarried from limestone beds that were formed by a very large foraminiferan population that flourished during the early Tertiary period.
Since fossil foraminiferans and radiolarians can be found in well drillings, their identification is often important to oil geologists for correlation of rock strata.
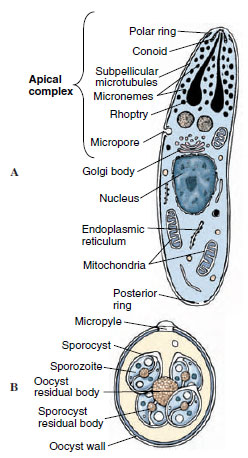 |
| Figure 11-19 A, Diagram of an apicomplexan sporozoite or merozoite at the electron microscope level, illustrating the apical complex. The polar ring, conoid, micronemes, rhoptries, subpellicular microtubules, and micropore (cytostome) are all considered components of the apical complex. B, Infective oocyst of Eimeria. The oocyst is the resistant stage and has undergone multiple fission after zygote formation (sporogony). |
Phylum Apicomplexa
All apicomplexans are endoparasites, and their hosts are found in many animal phyla. The presence of a certain combination of organelles, the apical complex, distinguishes this phylum (Figure 11-19A). The apical complex is usually present only in certain developmental stages of the organisms; for example, merozoites and sporozoites (Figure 11-20). Some structures, especially the rhoptries and micronemes, apparently aid in penetrating the host’s cells or tissues.
Locomotor organelles are less obvious in this group than in other protozoa. Pseudopodia occur in some intracellular stages, and gametes of some species are flagellated. Tiny contractile fibrils can form waves of contraction across the body surfaces to propel the organism through a liquid medium.
The life cycle usually includes both asexual and sexual reproduction, and sometimes an invertebrate intermediate host. At some point in the life cycle, the organisms develop a spore (oocyst), which is infective for the next host and is often protected by a resistant coat.
Class Sporozoa
The most important class of phylum Apicomplexa, Sporozoa, contains three subclasses: Gregarinia, Coccidia, and Piroplasmia. Gregarines are common parasites of invertebrates, but they are of little economic significance. Piroplasms are of some veterinary importance; for example, Babesia bigemina causes Texas red-water fever in cattle. Humans are occasionally infected with species of Babesia normally parasitic in other animals.
Subclass Coccidia Coccidia are intracellular parasites in invertebrates and vertebrates, and the group includes species of very great medical and veterinary importance.
Eimeria. The name “coccidiosis” is generally applied only to infections with Eimeria or Isospora. Humans can be infected with species of Isospora, but there is usually little disease. However, Isospora infections can be very serious in AIDS patients. Some species of Eimeria may cause serious disease in some domestic animals. Symptoms usually include severe diarrhea or dysentery.
Eimeria tenella is often fatal to young fowl, producing severe pathogenesis in the intestine. The organisms undergo schizogony in the intestinal cells, finally producing gametes. After fertilization the zygote forms an oocyst that passes out of its host in the feces (Figure 11-19B). Sporogony occurs within the oocyst outside the host, producing eight sporozoites in each oocyst. Infection occurs when a new host accidentally ingests a sporulated oocyst and the sporozoites are released by digestive enzymes.
Toxoplasma gondii. A similar life cycle occurs in Toxoplasma gondii, a parasite of cats, but this species produces extraintestinal stages as well. When rodents, cattle, sheep, humans, many other mammals, or even birds, ingest sporozoites, the sporozoites cross from the intestine and begin rapid, asexual reproduction in a variety of tissues. As the host mounts an immune response (see Immunity), reproduction of the zoites slows, and they become enclosed in tough tissue cysts. The zoites, now called bradyzoites, accumulate in large numbers in each tissue cyst. Bradyzoites are infective for other hosts, including cats, where they can initiate the intestinal cycle in a cat that eats infected prey. Bradyzoites can remain viable and infective for months or years, and it is estimated that onethird of the world’s human population carries tissue cysts containing bradyzoites in their body. Up to 50% of the human population of the United States are infected. The normal route of infection for humans is apparently consumption of infected meat that is insufficiently cooked.
In humans Toxoplasma causes little or no ill effects except in AIDS patients or in women infected during pregnancy, particularly in the first trimester. Such infection greatly increases the chances of a birth defect in the baby; it is now believed that 2% of all mental retardation in the United States is a result of congenital toxoplasmosis. Toxoplasmosis can also be a serious disease in persons who are immunosuppressed, either with drugs or by AIDS. In such patients rupture of a tissue cyst, which would be contained easily in a person with a normal immune system, becomes a source of life-threatening infection.
Plasmodium: The Malarial Organism: The best known of the coccidians is Plasmodium spp., causative organisms of the most important infectious disease of humans: malaria. Malaria is a very serious disease, difficult to control and widespread, particularly in tropical and subtropical countries. Four species of Plasmodium infect humans. Although each species produces its own peculiar clinical picture, all four have similar cycles of development in their hosts (Figure 11-20).
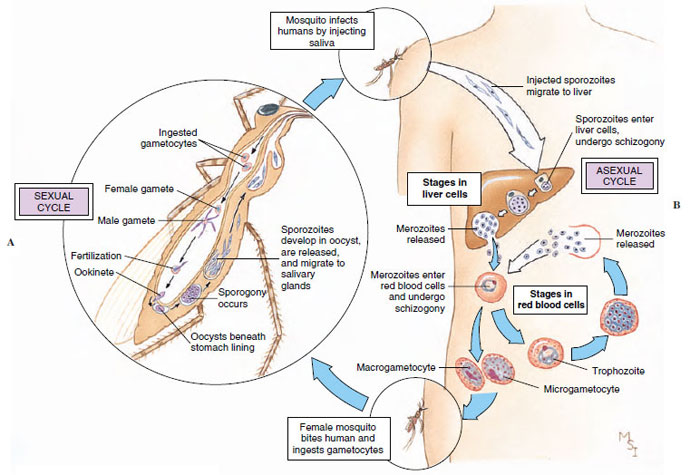 |
| Figure 11-20 Life cycle of Plasmodium vivax, one of the protozoa (class Sporozoa) that causes malaria in humans. A, Sexual cycle produces sporozoites in body of mosquito. Meiosis occurs just after zygote formation (zygotic meiosis). B, Sporozoites infect a human and reproduce asexually, first in liver cells and then in red blood cells. Malaria is spread by Anopheles mosquito, which ingests gametocytes along with human blood, then, when biting another victim, leaves sporozoites in new wound. |
The parasite is carried by mosquitoes (Anopheles), and sporozoites are injected into a human with the insect’s saliva during its bite. Sporozoites penetrate liver cells and initiate schizogony. The products of this division then enter other liver cells to repeat the schizogonous cycle, or in P. falciparum they penetrate the red blood cells after only one cycle in the liver. The period when the parasites are in the liver is the incubation period, and it lasts from 6 to 15 days, depending on the species of Plasmodium.
Merozoites released as a result of liver schizogony enter red blood cells, where they begin a series of schizogonous cycles. When they enter red blood cells, they become ameboid trophozoites, feeding on hemoglobin. The end product of the parasite’s digestion of hemoglobin is a dark, insoluble pigment: hemozoin. Hemozoin accumulates in the host cell, is released when the next generation of merozoites is produced, and eventually accumulates in the liver, spleen, or other organs. The trophozoite within a red blood cell grows and undergoes schizogony, producing 6 to 36 merozoites, which, depending on the species, burst forth to infect new red cells. When a red blood cell containing merozoites bursts, it releases the parasite’s metabolic products, which have accumulated there. Release of these foreign substances into the patient’s circulation results in the chills and fever characteristic of malaria.
Since the populations of schizonts maturing in red blood cells are synchronized to some degree, the episodes of chills and fever have a periodicity characteristic of the particular species of Plasmodium. In P. vivax (benign tertian) malaria and P. ovale malaria, episodes occur every 48 hours; in P. malariae (quartan) malaria, every 72 hours; and in P. falciparum (malignant tertian) malaria, about every 48 hours, although synchrony is less well defined in this species. People usually recover from infections with the first three species, but mortality may be high in untreated cases of P. falciparum infection. Sometimes grave complications, such as cerebral malaria, occur. Unfortunately, P. falciparum is the most common species, accounting for 50% of all malaria in the world. Certain genes, for example the gene for sickle cell hemoglobin, confer some resistance to malaria on people that carry them.
After some cycles of schizogony in red blood cells, infection of new cells by some of the merozoites results in production of microgametocytes and macrogametocytes rather than another generation of merozoites. When gametocytes are ingested by a mosquito feeding on a patient’s blood, they mature into gametes, and fertilization occurs. The zygote becomes a motile ookinete, which penetrates the stomach wall of the mosquito and becomes an oocyst. Within the oocyst, sporogony occurs, and thousands of sporozoites are produced. The oocyst ruptures, and the sporozoites migrate to the salivary glands, from which they are transferred to a human by a bite of the mosquito. Development in a mosquito requires 7 to 18 days but may be longer in cool weather.
Elimination of mosquitoes and their breeding places by insecticides, drainage, and other methods has been effective in controlling malaria in some areas. However, difficulties in carrying out such activities in remote areas and areas suffering civil unrest, and acquisition of resistance to insecticides by mosquitoes and to antimalarial drugs by Plasmodium (especially P. falciparum), mean that malaria will be a serious disease of humans for a long time to come.
Other species of Plasmodium parasitize birds, reptiles, and mammals. Those of birds are transmitted chiefly by Culex mosquitoes.
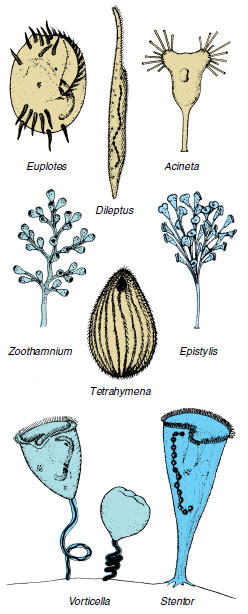 |
| Figure 11-21 Some representative ciliates. Euplotes have stiff cirri used for crawling about. Contractile fibrils in ectoplasm of Stentor and in stalks of Vorticella allow great expansion and contraction. Note the macronuclei, long and curved in Euplotes and Vorticella, shaped like a string of beads in Stentor. |
Phylum Ciliophora
Ciliates are a large and interesting group, with a great variety of forms living in all types of freshwater and marine habitats. They are the most structurally complex and diversely specialized of all protozoan groups. The majority are free living, but some are commensal or parasitic. They are usually solitary and motile, but some are sessile and some colonial. There is great diversity of shape and size. In general, they are larger than most other protozoa, but they range from very small (10 to 12 µm) up to 3 mm long. All have cilia that beat in a coordinated rhythmical manner, although the arrangement of the cilia may vary and some lack cilia as adults.
Ciliates are always multinucleate, possessing at least one macronucleus and one micronucleus, but varying from one to many of either type. The macronuclei are apparently responsible for metabolic and developmental functions and for maintaining all the visible traits, such as the pellicular apparatus. Macronuclei vary in shape among the different species (Figures 11-21 and 11-23). Micronuclei participate in sexual reproduction and give rise to macronuclei after exchange of micronuclear material between individuals. Micronuclei divide mitotically, and macronuclei divide amitotically.
The pellicle of ciliates may consist only of a cell membrane or in some species may form a thickened armor. Cilia are short and usually arranged in longitudinal or diagonal rows. Cilia may cover the surface of the organism or may be restricted to the oral region or to certain bands. In some forms cilia are fused into a sheet called an undulating membrane or into smaller membranelles, both used to propel food into the cytopharynx (gullet). In other forms there may be fused cilia forming stiffened tufts called cirri, often used in locomotion by the creeping ciliates (Figure 11-21).
An apparently structural system of fibers, in addition to the kinetosomes, makes up the infraciliature, just beneath the pellicle (Figure 11-22). Each cilium terminates beneath the pellicle in its kinetosome, and from each kinetosome a fibril arises and passes along beneath the row of cilia, joining with the other fibrils of that row. The cilia, kinetosomes, and other fibrils of that ciliary row make up what is called a kinety (Figure 11-22). All ciliates seem to have kinety systems, even those that lack cilia at some stage. The infraciliature apparently does not coordinate ciliary beat, as formerly thought. Coordination of ciliary movement seems to be by waves of depolarization of the cell membrane moving down the organism, similar to the phenomenon in a nerve impulse.
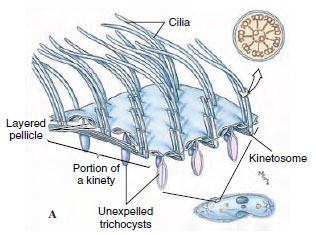 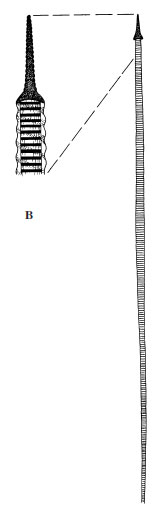 |
| Figure 11-22 Infraciliature and associated structures in ciliates. A, Structure of the pellicle and its relation to the infraciliature system. B, Expelled trichocyst. |
Most ciliates are holozoic. Most of them possess a cytostome (mouth) that in some forms is a simple opening and in others is connected to a gullet or ciliated groove. The mouth in some is strengthened with stiff, rodlike trichites for swallowing larger prey; in others, such as paramecia, ciliary water currents carry microscopic food particles toward the mouth. Didinium has a proboscis for engulfing paramecia on which it feeds (Figure 11-9). Suctorians paralyze their prey and then ingest the contents through tubelike tentacles by a complex feeding mechanism that apparently combines phagocytosis with a sliding filament action of microtubules in the tentacles (Figure 11-9).
Some ciliates have curious small bodies in their ectoplasm between the bases of the cilia. Examples are trichocysts (Figure 11-22 and 11-23) and toxicysts. Upon mechanical or chemical stimulation, these bodies explosively expel a long, threadlike structure. The mechanism of expulsion is unknown. The function of trichocysts is thought to be defensive. When attacked by a Didinium, a paramecium expels its trichocysts but to no avail. Toxicysts, however, release a poison that paralyzes the prey of carnivorous ciliates. Toxicysts are structurally quite distinct from trichocysts. Many dinoflagellates have structures very similar to trichocysts.
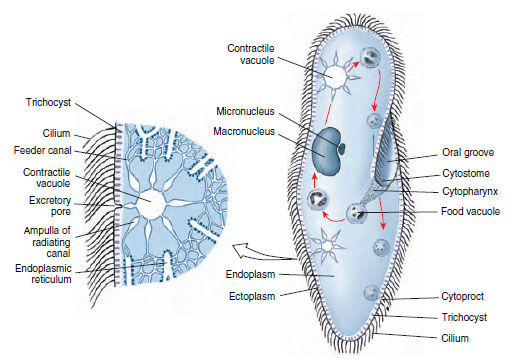 |
| Figure 11-23 Left, enlarged section of a contractile vacuole (water expulsion vesicle) of Paramecium. Water is apparently collected by endoplasmic reticulum, emptied into feeder canals and then into the vesicle. The vesicle contracts to empty its contents to the outside, thus serving as an osmoregulatory organelle. Right, Paramecium, showing cytopharynx, food vacuoles, and nuclei. |
Among the more striking and familiar of ciliates are Stentor (Gr. herald with a loud voice), trumpet shaped and solitary, with a beadshaped macronucleus (Figure 11-21); Vorticella (L. dim. of vortex, a whirlpool), bell shaped and attached by a contractile stalk (Figure 11-21); and Euplotes (Gr. eu, true, good, + ploter, swimmer) with a flattened body and groups of fused cilia (cirri) that function as legs.
Paramecium: A Representative Ciliate
Paramecia are usually abundant in ponds or sluggish streams containing aquatic plants and decaying organic matter.
Form and Function Paramecia are often described as slipper shaped. Paramecium caudatum is 150 to 300 µm in length and is blunt anteriorly and somewhat pointed posteriorly (Figure 11-23). The organism has an asymmetrical appearance because of the oral groove, a depression that runs obliquely backward on the ventral side.
The pellicle is a clear, elastic membrane that may be ornamented by ridges or papillalike projections (Figure 11-22), and its entire surface is covered with cilia arranged in lengthwise rows. Just below the pellicle is the thin clear ectoplasm that surrounds the larger mass of granular endoplasm (Figure 11-23). Embedded in ectoplasm just below the surface are spindleshaped trichocysts, which alternate with the bases of cilia. The infraciliature can be seen only with special fixing and staining methods.
The cytostome at the end of the oral groove leads into a tubular cytopharynx, or gullet. Along the gullet an undulating membrane of modified cilia keeps food moving. Fecal material is discharged through a cytoproct posterior to the oral groove (Figure 11-23). Within the endoplasm are food vacuoles containing food in various stages of digestion. There are two contractile vacuoles, each consisting of a central space surrounded by several radiating canals (Figure 11-23) that collect fluid and empty it into the central vacuole.
Paramecium caudatum has two nuclei: a large kidney-shaped macronucleus and a smaller micronucleus fitted into the depression of the former. These can usually be seen only in stained specimens. The number of micronuclei varies in different species; for example, P. multimicronucleatum may have as many as seven.
Paramecia are holozoic, living on bacteria, algae, and other small organisms. Cilia in the oral groove sweep food particles in the water into the cytostome, from which point they are carried into the cytopharynx by the undulating membrane. From the cytopharynx food is collected into a food vacuole that is constricted into the endoplasm. Food vacuoles circulate in a definite course through the cytoplasm while the food is being digested by enzymes from the endoplasm. The indigestible part of the food is ejected through the cytoproct.
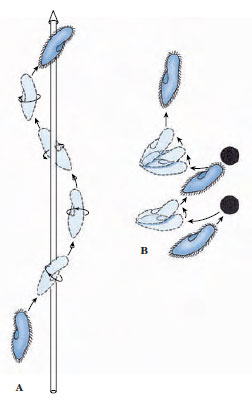 |
| Figure 11-24 A, Spiral path of swimming Paramecium. B, Avoidance reaction of Paramecium. |
The body is elastic, allowing it to bend and squeeze its way through narrow places. Its cilia can beat either forward or backward, so that the organism can swim in either direction. The cilia beat obliquely, causing the organism to rotate on its long axis. In the oral groove the cilia are longer and beat more vigorously than the others so that the anterior end swerves aborally. As a result of these factors, the organism moves forward in a spiral path (Figure 11-24A).
When a ciliate, such as a paramecium, comes in contact with a barrier or a disturbing chemical stimulus, it reverses its cilia, backs up a short distance, and swerves the anterior end as it pivots on its posterior end. This behavior is called an avoiding reaction (Figure 11-24B). A paramecium may continue to change its direction to keep itself away from the noxious stimulus, and it may react in a similar fashion to keep itself within the zone of an attractant. A paramecium may also change its swimming speed. How does a paramecium “know” when to change directions or swimming speed? Interestingly, the reactions of the organism depend on the effects of the stimulus on the electrical potential difference across its cell membrane. Paramecia slightly hyperpolarize in attractants and depolarize in repellents that produce the avoiding reaction. Hyperpolarization increases the rate of the forward ciliary beat, and depolarization results in ciliary reversal and backward swimming.
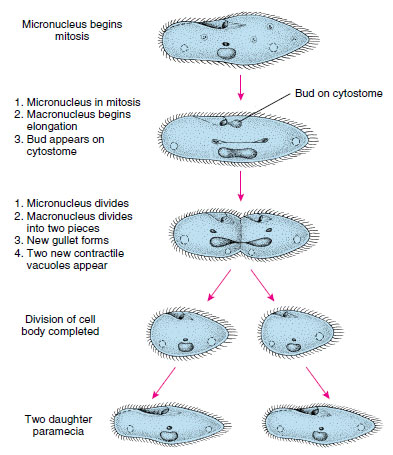
Reproduction Paramecia reproduce only by binary fission across kineties (ciliary rows) but have certain forms of sexual phenomena called conjugation and autogamy.
In binary fission the micronucleus divides mitotically into two daughter micronuclei, which move to opposite ends of the cell (Figure 11-25). The macronucleus elongates and divides amitotically.
 |
| Figure 11-25 Binary fission in a ciliophoran (Paramecium). Division is across rows of cilia. |
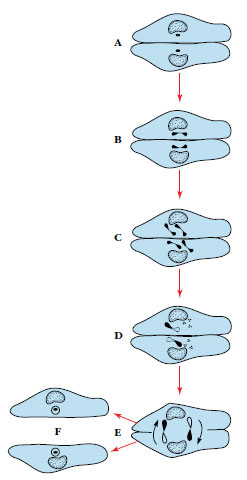 |
| Figure 11-26 Scheme of conjugation in Paramecium. A, Two individuals come in contact on their oral surface. B and C, Micronuclei divide twice (meiosis), resulting in four haploid nuclei in each partner. D, Three micronuclei degenerate; remaining one divides to form “male” and “female” pronuclei. E, Male pronuclei are exchanged between conjugants. F, Male and female pronuclei fuse, and individuals separate. Subsequently old macronuclei are absorbed and replaced by new macronuclei. |
The result of conjugation is similar to that of zygote formation, for each exconjugant contains hereditary material from two individuals. The advantage of sexual reproduction is that it permits gene recombinations, thus increasing genetic variation in the population. Although ciliates in clone cultures can apparently reproduce repeatedly and indefinitely without conjugation, the stock seems eventually to lose vigor. Conjugation restores vitality to a stock. Seasonal changes or a deteriorating environment will usually stimulate sexual reproduction.
Autogamy is a process of selffertilization that is similar to conjugation except that there is no exchange of nuclei. After the disintegration of the macronucleus and the meiotic divisions of the micronucleus, two haploid pronuclei fuse to form a synkaryon that is completely homozygous (Principles of Genetics:A Review).
Symbiotic Ciliates
Many symbiotic ciliates live as commensals, but some can be harmful to their hosts. Balantidium coli lives in the large intestine of humans, pigs, rats, and many other mammals (Figure 11-27). There seem to be hostspecific strains, and the organism is not easily transmitted from one species to another. Transmission is by fecal contamination of food or water. Usually the organisms are not pathogenic, but in humans they sometimes invade the intestinal lining and cause a dysentery similar to that caused by Entamoeba histolytica. The disease can be serious and even fatal. Infections are common in parts of Europe, Asia, and Africa but are rare in the United States.
Other species of ciliates live in other hosts. Entodinium (Figure 11-27) belongs to a group that has very complex structure and lives in the digestive tract of ruminants, where they may be very abundant. Nyctotherus live in the colon of frogs and toads. In aquarium and wild freshwater fishes, Ichthyophthirius causes a disease known to many fish culturists as “ick.” Untreated, it can cause much loss of exotic fishes.
Suctorians
Suctorians are ciliates in which the young possess cilia and are free swimming, and the adults grow a stalk for attachment, become sessile, and lose their cilia. They have no cytostome but feed by long, slender, tubelike tentacles. The suctorian captures living prey, usually a ciliate, by the tip of one or more tentacles and paralyzes it. The cytoplasm of the prey then flows through the attached tentacles, forming food vacuoles in the feeding suctorian (Figure 11-9).
One of the best places to find freshwater suctorians is in algae that grow on the carapace of turtles. Common genera of suctorians found there are Anarma (without stalk or test) and Squalorophrya (with stalk and test). Other freshwater representatives are Podophrya (Figure 11-9) and Dendrosoma. Acinetopsis and Ephelota are saltwater forms. Suctorian parasites include Trichophrya, the species of which are found on a variety of invertebrates and freshwater fish; Allantosoma, which live in the intestine of certain mammals; and Sphaerophrya, which are found in Stentor.
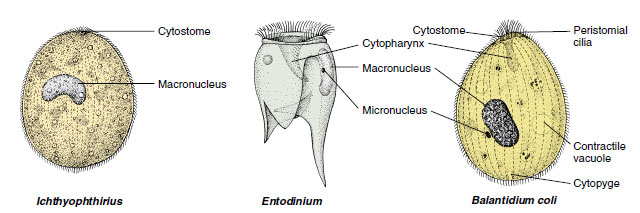 |
| Figure 11-27 Some symbiotic ciliates. Balantidium coli is a parasite of humans and other mammals. Ichthyophthirius causes a common disease in aquarium and wild freshwater fishes. Entodinium is found in the rumen of cows and sheep. |
Support our developers




