The Chordates
 |
| Two amphioxus in feeding posture. |
Along the more southern coasts of North America, half buried in sand on the sea floor, lives a small fishlike translucent animal quietly filtering organic particles from seawater. Inconspicuous, of no commercial value and largely unknown, this creature is nonetheless one of the famous animals of classical zoology. It is amphioxus, an animal that wonderfully exhibits the four distinctive hallmarks of the phylum Chordata—(1) dorsal, tubular nerve cord overlying (2) a supportive notochord, (3) pharyngeal slits for filter feeding, and (4) a postanal tail for propulsion—all wrapped up in one creature with textbook simplicity. Amphioxus is an animal that might have been designed by a zoologist for the classroom. During the nineteenth century, with interest in vertebrate ancestry running high, amphioxus was considered by many to resemble closely the direct ancestor of the vertebrates. Its exalted position was later acknowledged by Philip Pope in a poem sung to the tune of “Tipperary.” It ends with the refrain:
-
It’s a long way from amphioxus
-
It’s a long way to us.
-
To the meanest human cuss.
-
And its welcome lungs and hair,
-
But we all came from there.
Position in the Animal Kingdom
Phylum Chordata (kor-da´ta) (L. chorda, cord) belongs to the Deuterostomia branch of the animal kingdom that includes the phyla Echinodermata, Hemichordata, and the three lophophorate phyla—Phoronida, Ectoprocta, and Brachiopoda. These six phyla share many embryological features and are probably descended from an ancient common ancestor. From humble beginnings, the chordates have evolved a vertebrate body plan of enormous adaptability that always remains distinctive, while it provides almost unlimited scope for specialization in life habitat, form, and function.
Biological Contributions
- The endoskeleton of vertebrates permits continuous growth without molting and attainment of large body size, and it provides an efficient framework for muscle attachment.
- The perforated pharynx of protochordates that originated as a suspension-feeding device served as the framework for subsequent evolution of true internal gills with pharyngeal muscular pump, and jaws.
- Adoption of a predatory habit by the early vertebrates and accompanying evolution of a highly differentiated brain and paired special sense organs contributed in large measure to the successful adaptive radiation of vertebrates.
- Paired appendages that appeared in the aquatic vertebrates were successfully adapted later as jointed limbs for efficient locomotion on land or as wings for flight.
Characteristics of Phylum Chordata
- Bilateral symmetry; segmented body; three germ layers; welldeveloped coelom
- Notochord (a skeletal rod) present at some stage in the life cycle
- Single, dorsal, tubular nerve cord; anterior end of cord usually enlarged to form brain
- Pharyngeal pouches present at some stage in the life cycle; in aquatic chordates these develop into pharyngeal slits
- Postanal tail, usually projecting beyond the anus at some stage but may or may not persist
- Segmented muscles in an unsegmented trunk
- Ventral heart, with dorsal and ventral blood vessels; closed blood system
- Complete digestive system
- A cartilaginous or bony endoskeleton present in the majority of members (vertebrates)
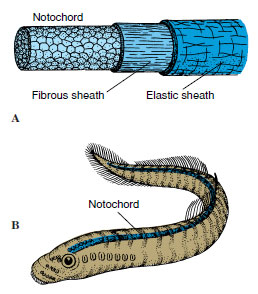 |
| Figure 25-1 A, Structure of the notochord and its surrounding sheaths. Cells of the otochord proper are thick walled, pressed together closely, and filled with semifluid. Stiffness is caused mainly by turgidity of fluid-filled cells and surrounding connective tissue sheaths. This primitive type of endoskeleton is characteristic of all chordates at some stage of the life cycle. The notochord provides longitudinal stiffening of the main body axis, a base for trunk muscles, and an axis around which the vertebral column develops. B, In hagfishes and lampreys it persists throughout life, but in other vertebrates it is largely replaced by vertebrae. In mammals slight remnants are found in nuclei pulposi of intervertebral discs. The method of notochord formation is different in the various groups of animals. In amphioxus it originates from endoderm; in birds and mammals it arises as an anterior outgrowth of the embryonic primitive streak. |
The Chordates
The animals most familiar to most people belong to the phylum Chordata (kor-da´ta) (L. chorda, cord). Humans are members and share with other chordates the characteristic from which the phylum derives its name—the notochord (Gr. noton, back, + L. chorda, cord) (Figure 25-1). All members of the phylum possess this structure, either restricted to early development or present throughout life. The notochord is a rodlike, semirigid body of cells enclosed by a fibrous sheath, which extends, in most cases, the length of the body between the gut tract and central nervous system. Its primary purpose is to support and stiffen the body, that is, to act as a skeletal axis.
The structural plan of chordates shares features of many nonchordate invertebrates, such as bilateral symmetry, anteroposterior axis, coelom, tube-within-a-tube arrangement, metamerism, and cephalization. However, the exact phylogenetic position of the chordates within the animal kingdom is unclear.
Two possible lines of descent have been proposed. Earlier speculations that focused on the arthropod-annelidmollusc group (Protostomia branch) of the invertebrates have fallen from favor. It is now believed that only members of the echinodermhemichordate assemblage (Deuterostomia branch) deserve serious consideration as a chordate sister group. Chordates share with the other Deuterostomes several important characteristics: radial cleavage, anus derived from the first embryonic opening (blastopore) and mouth derived from an opening of secondary origin, and a coelom primitively formed by fusion of enterocoelous pouches (except in vertebrates in which the coelom is basically schizocoelous). These common characteristics indicate a natural unity among the Deuterostomia.
As a whole, there is more fundamental unity of plan throughout all the organs and systems of this phylum than there is in many other phyla. Ecologically the chordates are among the most adaptable of organic forms and are able to occupy most kinds of habitat. They illustrate perhaps better than any other animal group the basic evolutionary processes of the origin of new structures, adaptive strategies, and adaptive radiation.
Traditional and Cladistic Classification of the Chordates
The traditional Linnaean classification of the chordates provides a simple and convenient way to indicate the taxa included in each major group. However, in cladistic usage, some of the traditional taxa, such as Agnatha and Reptilia, are no longer recognized. Such taxa do not satisfy the requirement of cladistics that only monophyletic groups are valid taxonomic entities, that is, groups that contain all known descendants of a single common ancestor. The reptiles, for example, are considered a paraphyletic grouping because this group does not contain all of the descendants of their most recent common ancestor. The common ancestor of reptiles as traditionally recognized is also the ancestor of birds and mammals. As shown in the cladogram (Figure 25-3), reptiles, birds, and mammals compose a monophyletic clade called Amniota, so named because all develop from an egg having special extraembryonic membranes, one of which is the amnion. Therefore according to cladistics, the reptiles can be grouped only in a negative manner as amniotes that are not birds or mammals; there are no positive or novel features that unite reptiles to the exclusion of birds and mammals. Similarly, agnathans (hagfishes and lampreys) are a paraphyletic grouping because the most recent common ancestor of agnathans is also an ancestor of all remaining vertebrates (the gnathostomes). The reasons why paraphyletic groups are not used in cladistic taxonomy are explained in Classification and Phylogeny of Animals.
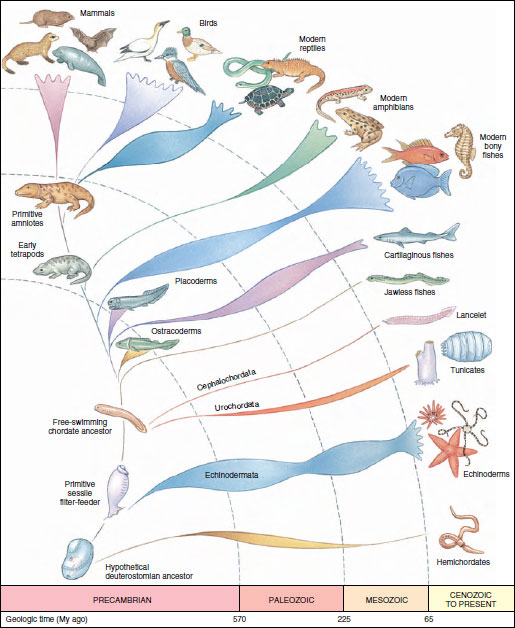 |
| Figure 25-2 Phylogenetic tree of the chordates, suggesting probable origin and relationships. Other schemes have been suggested and are possible. The relative abundance in numbers of species of each group through geological time, as indicated by the fossil record, is suggested by the bulging and thinning of that group’s line of descent. |
The phylogenetic tree of the chordates (Figure 25-2) and the cladogram of the chordates (Figure 25-3) provide different kinds of information. The cladogram shows a nested hierarchy of taxa grouped by their sharing of derived characters. These characters may be morphological, physiological, embryological, behavioral, chromosomal, or molecular in nature. Although the cladogram shows the relative time of origin of the novel properties of taxonomic groups and their specific positions in the hierarchical system of evolutionary common descent, it contains no timescale or information on ancestral lineages. By contrast, the branches of a phylogenetic tree are intended to represent real lineages that occurred in the evolutionary past. Geological information regarding ages of lineages is added to information from the cladogram to generate a phylogenetic tree for the same taxa.
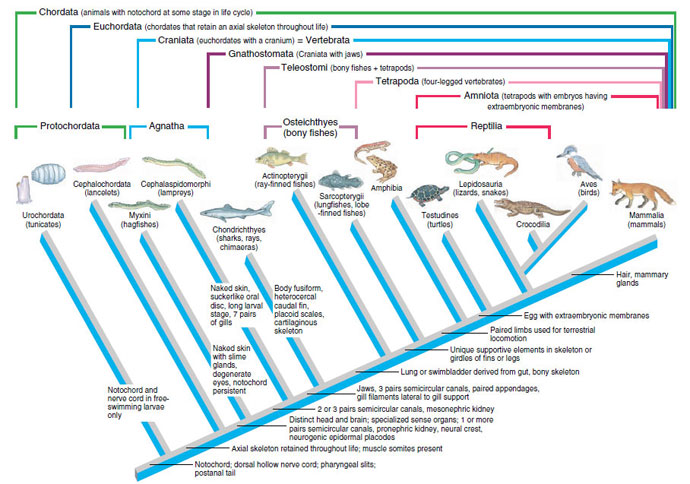 |
| Figure 25-3 Cladogram of living members of phylum Chordata showing probable relationships of monophyletic groups composing the phylum. Each branch in the cladogram represents a monophyletic group. Some derived character states that identify the branchings are shown at right of the branch points. Nesting brackets across the top of the cladogram identify monophyletic groupings within the phylum. The term Craniata, although commonly equated with Vertebrata, is preferred by many authorities because it recognizes that jawless vertebrates (Agnatha) have a cranium but no vertebrae. The lower set of brackets identify the traditional groupings Protochordata, Agnatha, Osteichthyes, and Reptilia. These paraphyletic groups are not recognized in cladistic treatments, but are shown because of widespread use. |
In our treatment of the chordates, we have retained the traditional Linnaean classification because of its conceptual usefulness and because the alternative—thorough revision following cladistic principles— would require extensive change and virtual abandonment of familiar rankings. However, we have tried to use monophyletic taxa as much as possible, because such usage is consistent with both evolutionary and cladistic taxonomy.
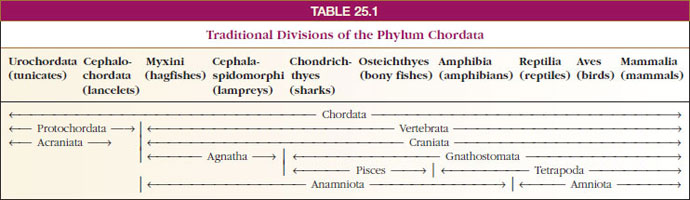
Several traditional divisions of the phylum Chordata used in Linnaean classifications are shown in Table 25-1. A fundamental separation is Protochordata from Vertebrata. Since the former lack a well-developed head, they are also called Acraniata. All vertebrates have a well-developed skull case enclosing the brain and are called Craniata. The vertebrates (craniates) may be variously subdivided into groups based on shared possession of characteristics. Two such subdivisions shown in Table 25-1 are: (1) Agnatha, vertebrates lacking jaws (hagfishes and lampreys), and Gnathostomata, vertebrates having jaws (all other vertebrates) and (2) Amniota, vertebrates whose embryos develop within a fluid-filled sac, the amnion (reptiles, birds, and mammals), and Anamniota, vertebrates lacking this adaptation (fishes and amphibians). The Gnathostomata in turn can be subdivided into Pisces, jawed vertebrates with limbs (if any) in the shape of fins; and Tetrapoda (Gr. tetras, four, + podos, foot), jawed vertebrates with two pairs of limbs. Note that several of these groupings are paraphyletic (Protochordata, Acraniata, Agnatha, Anamniota, Pisces) and consequently are not accepted in cladistic classifications. Accepted monophyletic taxa are shown at the top of the cladogram in Figure 25-3 as a nested hierarchy of increasingly more inclusive groupings.
 |
Support our developers




