Several hypothesis were put forward to explain inbreeding depression and conversely to explain the related phenomenon of hybrid vigour. Two of them, proposed within the first decade after rediscovery of Mendel's work, claimed wide acceptance. These were
'dominance' and
'overdominance' hypotheses. These will be discussed in some detail in the following text.
Dominance hypothesis
The hypothesis, first proposed in 1908 by
Davenport and later in 1910 supported independently by
Bruce and by
Keeble and
Pellew assumes that a cross fertilizing species consists of a large number of genetically different individuals, many of which carry deleterious recessive genes concealed in heterozygotes. Upon inbreeding in such individuals, various morbid homozygous recessive types appear.
These include chlorophyll deficiency, malformation in flowers, defective seeds, etc., which inhibit reproduction thus eliminating them. Still others have some mild characteristics like partial chlorophyll deficiency, dwarfness and deficient root system not leading to direct extinction, but nevertheless the development is severely handicapped. In open- pollinated species also, these characters appear, though in much lower frequency. Unfavourable genes segregate on inbreeding, and, upon fixation brought by homozygosity, produce lines that possess different gene complexes. Some lines receive more favourable genes than others, accounting for differences observed in the degree of inbreeding depression in different lines. Thus, it is quite clear that the inbreeding depression is not a process of degeneration but a consequence of Mendelian segregation.
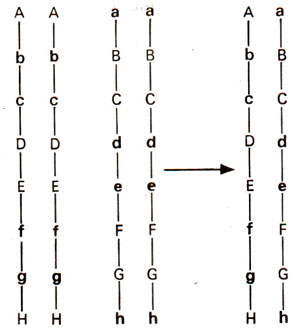
Fig. 47.3. A hypothetical cross between inbred lines homozygous for different deleterious recessives on a particular chromosome leading to a heterotic hybrid. The chromosome pair in the hybrid has all the deleterious recessive genes of both the parents but also bears normal dominant gene for each recessive.
According to this hypothesis, the intercrossing of inbreds will give hybrids having deleterious recessives of one parent, hidden again, by dominant alleles contributed by the other parent, as in the original open pollinated stock. It seems, therefore, that the precise degree of response to crossing should be a function of the genotypes of particular inbreds. Some genotypes should complement each other nicely to produce hybrids better than the average of the original open-pollinated variety, whereas others will not. This is due to specific combinations of dominant and recessive genes they happened to receive during segregation.
There were two main objections against this hypothesis.
First, if the hypothesis is correct, it should be possible to obtain individuals homozygous for all the dominant genes. Such lines as vigorous as F
1 should be true breeding. However, such high yielding homozygous lines could not be recovered.
Jones (1917) proposed
'linkage of dominant growth factors' to counter this objection. According to him probably many genes affect growth and that each chromosome would be expected to contain several of these genes.
A single linkage group would be having some unfavourable recessives in addition to favourable dominants. Some of these combinations would pass from one generation to the next without free recombinations, and homozygous lines as vigorous as F
1 hybrid may not be obtained. A series of precisely placed crossovers would be required to obtain all the dominant alleles in one gamete. This can be illustrated using the Figure 47.3. If we consider the genes A to H on a single chromosome, crossing over must take place between A and B, C and D, E and F, G and H to produce a gamete with dominant alleles at all loci. Crossing over at less than all of these points or at regions other than those given above along the chromosome (i.e. B and C, D and E, F and G) would be inadequate for obtaining all dominant alleles.
Fig. 47.3. A hypothetical cross between inbred lines homozygous for different deleterious recessives on a particular chromosome leading to a heterotic hybrid. The chromosome pair in the hybrid has all the deleterious recessive genes of both the parents but also bears normal dominant gene for each recessive.
The
second objection to this theory was that if heterosis were solely due to dominance of independent factors, the F
2 distribution curve should be skewed, because dominant and recessive phenotypes would be distributed according to the expansion of the binomial (3/4 + l/4)
n. To overcome this objection,
Jones suggested that due to linkage between groups of favourable and unfavourable genes it may not possible to obtain a highly skewed F
2 distribution.
Collins in 1921, however suggested that linkage is not necessarily the cause for the absence of skewness, because where a larger number of factors is involved, say more than ten, the chances of obtaining a single plant homozygous for all ten factors are very remote. As the number of factor pairs increases, the probability of the occurrence of completely homozygous dominant individuals decreases very rapidly.
There are also experiments supporting
Interallelic or
Dominant hypothesis. Quinby and
Karper (1946) studied heterosis in
Sorghum and observed that the heterozygote
Mama is significantly late in maturity and produces a greater weight of grain than either of the homozygote parents,
MaMa or
mama. Keeble and
Pellew studied two varieties of pea, both semi-dwarf, one with thin stem and long internodes, and the other with thick stems and short internodes. The F
1 hybrid was much taller than either of the parents, combining the long internodes of one parent with many nodes of the other.
Rickey reported a similar case in corn where dwarf x dwarf gave tall plants.
Crow (1940) attempted to calculate the maximum heterosis possible under the dominance hypothesis and concluded that if all the recessives were replaced by their dominant alleles, the vigour of an equilibrium population would be increased by 5%. This can be interpreted as the average maximum improvement in vigour, as measured in terms of selection advantage, that could occur because of hybridization.
Overdominance hypothesis
The overdominance hypothesis was proposed in 1908 by
Shull and
East, independently. It is assumed here that there is a physiological stimulus to development that increases with the diversity of the uniting gametes. In other words, using Mendelian terms, it means that there are loci at which the heterozygote is superior to either homozygote and that vigour increases in proportion to the amount of heterozygosity.
Various workers have given different names to this hypothesis such as
'single-gene heterosis', 'cumulative action of divergent alleles' and
'stimulations of divergent alleles'. Fisher (1930) called it
superdominance. In sirripler terms, this hypothesis assumes that the heterozygous combination
a1a2 at a single locus is superior to either of the homozygous combinations,
a1a1, or
a2a2. This implied that
a1and
a2perform different functions and the sum of their different products TS-sttpefioF to the single product produced by either allele in homozygous state. In 1936,
East further elaborated the idea by giving.
'Interallelic series of growth factors hypothesis', which proposes the presence of a series of alleles
a1,
a2, a3,
a4, ... of gradually increasing divergent functions. Heterozygotes were postulated to become increasingly more efficient, as their component alleles were more divergent, e.g.
a1a2 < a1a3 < a1a4 and so forth. There was no direct evidence of a locus at which heterozygote lies outside the range of the homozygotes, at that time.
However, now there are some evidences that different alleles at the same locus can do different things
(Whalay, 1952). For example, in maize, the heterozygote is more pigmented than either of the two homozygotes, in presence of certain R alleles. In animals, members of a multiple allelic series have been found that produce different blood antigens.
Gustafsson, while working with spontaneous mutants in pure lines of barley, reported two instances where heterozygotes for some chlorophyll mutations, produced more and larger seeds than homozygous normal plant. However, lack of clear-cut evidences of single gene heterosis has forbidden its general acceptance.
In most cases, both the hypotheses lead to same expectations. The difference is in the impossibility in obtaining homozygotes as vigorous as heterozygotes if single gene overdominance is important in heterosis. Evidences on this point are, however, difficult to obtain. Despite extensive work over the years, no conclusive evidence has come favouring one or the other hypothesis. However, there are reasons to believe that both systems can remain operative simultaneously in producing heterotic effect. The genes affecting heterosis can be expected to be as complex in their interaction as the genes in classical genetics. If this is true, we can say that both the hypotheses are gross oversimplification of the actual complex situation.




