Meiotic division consists of two successive divisions of a cell, so that as a result of one complete meiotic division, four cells will result. First division is accompanied with reduction in chromosome number without any division of chromosomes, while second division involves separation of chromatids of the chromosomes. Consequently, number of chromosomes which is reduced in first division remains constant (haploid) during second division. For convenience, therefore, meiosis can be described in two parts (i) first meiotic division, and (ii) second meiotic division.
Preceding meiosis, there is an interphase just like the one found in mitosis, consisting of G
1phase, S phase and G
2 phase. However in meiosis G
2 phase is very short or altogether absent, so that meiotic division takes over just after DNA synthesis is complete.
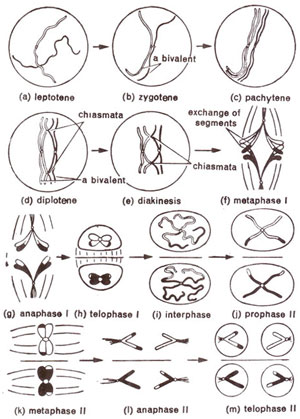
Fig. 7.10. Various stages of meiosis in a mother cell, (for clarity, chromosomes at different stages are not shown in their relative sizes).
First meiotic division.
First meiotic division is more important than second meiotic division, since it i& the reduction division, while second division is like a mitotic division. Like any other cell division, first meiotic division also consists of prophase, metaphase, anaphase and telophase.
(a) First prophase.
First prophase is of a very long duration and is also complex, differing from mitotic prophase in several respects. In order to describe it in detail, first prophase has been subdivided into five stages, namely, leptotene, zygotene, pachytene, diplotene and diakinesis. These five stages are established for convenience of description. Otherwise such a distinction is not possible and one stage very gradually passes on into the next stage.
(i) Leptotene (leptonema). This is the first stage of meiosis following interphase. Chromosomes at this stage appear as long thread-like structures, which are loosely interwoven. On these thread-like chromosomes, bead-like structures called chromomeres are found all along the length of chromosomes (Fig. 7.10a). In some animals, particularly in insects, chromosomes have their ends drawn together at a point on nuclear membrane, near centriole. Absence of such polarization of chromosome ends in plants is perhaps due to absence of centrosome. However, in certain plants like
Lilium, the chromosomes are densely clumped to one side, no chromatin material being seen in the rest of the nucleus. This phenomenon is called synizesis and its significance is not clearly understood.
Fig. 7.10. Various stages of meiosis in a mother cell, (for clarity, chromosomes at different stages are not shown in their relative sizes).
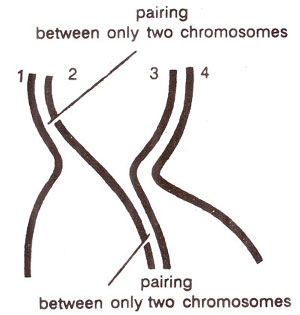
Fig 7.11. Meiotic pairing among four homologous chromosomes.
(ii) Zygotene (zygonema). Zygotene is characterized by pairing of homologous chromosomes (synapsis). The pairing is brought about in a zipper- like fashion (Fig. 7.10b) and may start at centromere, at chromosome ends or at any other position. This pairing takes place between homologous segments even if they are present in non-homologous chromosomes (e.g. in case of translocations).
Another important feature of this pairing is that pairing is allowed only between two chromosomes in one region. For instance, in an autotetraploid, where there will be four homologous chromosomes, in a particular region only two chromosomes willpair (Fig. 7.11). It
seems that as soon as two chromosomes come in contact, their attraction forces are neutralized. The only exception is the pairing between giant chromosomes in somatic cells of salivary glands of
Drosophila. In this case, if there are three or more homologous chromosomes, these will pair throughout their length, but this is not meiotic pairing.
Fig 7.11. Meiotic pairing among four homologous chromosomes.
An important question at this stage, is what are those factors which bring about pairing of chromosomes in meiotic prophase? This aspect has also been studied at the biochemical level.
Darlington (1930) gave an explanation for meiotic chromosome pairing in the form of
precocity theory.
According to this theory, meiosis is a precocious mitosis, as a result of which the chromosomes which have not yet duplicated, have to enter prophase. This is responsible for chromosome pairing. However, recent work has shown that DNA synthesis is completed before the onset of meiosis. Therefore, precocity theory is no longer tenable.
Stern and
Hotta (1969) have shown that premeiotic S-phase, unlike premitotic one, does not include complete chromosome replication. About 0.3% DNA, forming a significant component replicates only at about zygotene and is believed to control chromosome pairing (consult next main topic for details).
(iii) Pachytene (pachynema). Once homologous chromosomes have undergone pairing at zygotene, the cell enters the stage of pachytene, where chromosomes become shortened and coiled. The chromosomes appear as thickened thread-like structures, haploid in number (Fig. 7.10c). Each thread, however, if carefully examined has two homologous chromosomes closely appressed against each other. These pairs of homologous chromosomes are called bivalents. Each chromosome in a bivalent at this stage has two chromatids, as a result of which a bivalent really consists of four chromatids and is called a
tetrad. At this stage, crossing over or exchange of segments of chromatids is brought about. The nucleolus still persists. Attached to nucleolus can be seen the nucleolar organizing bivalent.
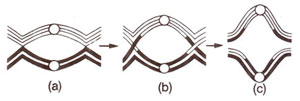
Fig. 7.12. Terminalization Qf chiasmata during prophase I.
(iv) Diplotene (diplonema). At diplotene, further thickening and shortening of chromosomes take place. Homologous chromosomes start separating from one another. This separation starts at centromeres, and travels towards the ends (Fig. 7.10d).
This kind of separation from centromere towards the ends is known as
terminalization. Due to such separation, dual nature of a bivalent becomes distinct and hence the name diplotene. Homologous chromosomes are now held together only at certain points along the length. Such points of contact between homologous chromosomes are known as
chiasmata (singular,
chiasma) and represent the places of crossing over (actual exchange of chromosome segments). As terminalization occurs, these chiasmata move towards the ends of chromosomes (Fig. 7.12). Number of chiasmata per bivalent is normally dependent on the length of chromosomes. These can be terminal when found at the end of bivalents due to terminalization effect or can be intercalary. The terminal chiasmata also originate at intercalary positions.
During crossing over at one point, only two chromatids, one from each of the two homologous chromosomes, take part. It should be emphasized that chiasmata are not the cause but are only the consequence of crossing over. There are various views explaining the mechanism of crossing over and chiasma formation. These views will be discussed in
Linkage and Crossing Over in Diploid Organisms (Higher Eukaryotes) and
Molecular Mechanisms of Recombination.
Fig. 7.12. Terminalization Qf chiasmata during prophase I.
(v) Diakinesis. At diakinesis, chromosomes continue to undergo further contraction. Ordinarily the only distinction between diplotene and diakinesis is the more contracted state of bivalents at diakinesis (Fig. 7.10e). Nucleolus may not be seen at this stage. Due to further terminalization and contraction, bivalents appear as rounded bodies darkly stained (with acetocarmine) and evenly scattered throughout the cell. Consequently, chiasmata are mainly terminal and chromosome counting is easy at this stage. In cases of longer chromosomes with intercalary chiasmata, all chiasmata are not terminalized, some of them still being in the intercalary region.
(b) First metaphase.
At metaphase, chromosomes are most condensed and have a smooth outline. The
spindle apparatus starts appearing and bivalents become attached to spindle through centromeres. Bivalents then appear in the form of an equatorial plate, due to the movement known as
congression (Fig. 7.10f).
Some workers describe a prometaphase stage between diakinesis and first metaphase. Whenever, it is described, it will include the period between the dissolution of the nuclear membrane and the full appearance of the spindle.
(c) First anaphase.
The movement of chromosomes of a bivalent from equatorial plate to poles constitute first anaphase. While in mitotic anaphase, centromere divides longitudinally and two sister chromatids pass to two different poles, in case of first anaphase of meiosis, sister chromatids do not separate but go to the same pole (Fig. 7.10g). When sister chromatids go to same pole, it is called a
reductional or
disjunctional division; on the other hand, when they separate and go to two poles it is an
equational division.
After anaphase I, each pole has a haploid number of chromosomes. Thus the chromosome number is reduced. The meiotic division is also called a reduction division, due to this reduction in chromosomes number. However, when we use the term reductional and equational to explain anaphase separation, we use it in the sense explained above.
(d) First telophase and interphase.
At first telophase, nuclear membranes are formed around the groups of chromosomes at the two poles (Fig. 7.10h). After formation of nuclei, chromosomes pass into a small interphase before the second meiotic division will start. In some cases, the first telophase and the subsequent interkinesis may be absent. In such cases,
chromosomes at the two poles after anaphase will directly pass to metaphase of the second division.
First meiotic division, which is completed at first telophase, may be followed by cytokinesis giving rise to a dyad (Fig. 7.10i). Such a division is called successive
division. However, cytokinesis may be postponed till the end of second division, when four cells are formed due to
simultaneous division.
Second meiotic division.
First meiotic division is followed by a second meiotic division with or without the intervening interphase. The second division is essentially a mitotic division and
is sometimes referred to as meiotic mitosis. Since mitosis has already been discussed, only a brief account of second meiotic division will be given here.
At second prophase, chromosomes are already double, each having two sister chromatids with a single functional centromere (Fig. 7.l0j). These chromosomes soon arrange at metaphase plate during second metaphase (Fig. 7.10k). The centromere, then splits and two chromatids, which may now be called chromosomes, pass to two poles during second anaphase (Fig. 7.10 1). This is soon followed by second telophase (Fig. 7.10m), and cytokinesis. As indicated earlier, cytokinesis may be successive or of simultaneous type and will give rise to four haploid cells.




