Palynology
» Pollen Morphology
» Polarity
» Symmetry
» Size
» Shape
» Aperture
» Position and Shape of apertures
» Polar apertures
» Global apertures
» Sporoderm (spore wall) structure
» Sexine patterns
» Excrescences
» Palynogram
» Palynology in relation of Honey Industry (Melittopalynology)
» Diagnosis of honeys
» Types of honey
» Geographical origin of honey
Studies in palynology began as early as the 18th century, but it came to be recognised as an authentic branch of study in 1916 at the Oslo conference. The study of pollen grains has gained significance in various fields of research, and the knowledge of pollen morphology has become an essential pre-requisite in applied palynology. The study of pollen morphology is also being made with a view to recognise and identify the fossil pollen microflora. Regional and local pollen flora, for the purpose of assisting the identification of the pollen fossils, has been prepared for different parts of the world by various scientists; a similar pollen flora has been prepared by Nair (1956) for the Western Himalayas in India.
In palynology, the pollen is mainly studied for pollen morphology, i.e., its polarity, symmetry, size, shape, aperture, and the sporoderm (exine) characters.
Pollen Morphology
The majority of the pollen grains have two coverings or walls called sporoderm. An outer layer is called exine or sclerine (sclerodermatous) while the inner is known as intine. The terms are also used for description of spores of mosses and ferns and instead of exine it’s called exosporium and for intine endosporium. When exine is formed of three layers, they are known as ektexine (the outermost layer of exine) mesexine (the middle layer) and endexine (the inner most layer of the exine — Fig. 12.1).
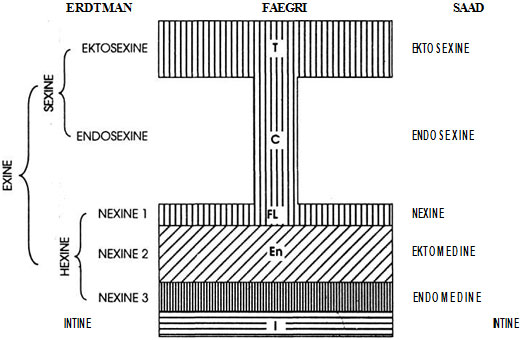 Fig. 12.1 Different terminology used by Palynologists to describe sporoderm
Fig. 12.1 Different terminology used by Palynologists to describe sporodermFaegri described the outer coat as exine, middle as endexine, and inner as intine. The three layers of exine are called tegillum the outermost, columella the middle and foot layer the innermost.
Saad described the outermost coat as exine, middle as medine and inner as intine. The two layers of exine are described as sexine and nexine. Nexine of Saad corresponds to foot layer of Faegri and nexine of Erdtman. He called tegillum as ektosexine and columella as endosexine. The two layers of medine are known as ektomedine and endomedine which correspond to nexine 2 and nexine 3 of Erdtman.
Polarity
The pollen grains (or spores) are produced in fours (tetrads) by spore mother cells. The polar axis of the pollen (spore) is the imaginary line passing through the centre of the spore to the centre of the tetrad (Fig. 12.2 A). Thus, in a tetrad, there can be four such lines. Each spore has two poles, the proximal and the distal. The proximal pole of a spore faces towards the centre of the tetrad and it is the proximal or inner face of the spore; the distal pole faces in the opposite direction, i.e., it is the outer face of the spore (Fig. 12.2 B). In case of normal spore tetrads, the spores are polar, i.e., their proximal and distal poles can be easily notable.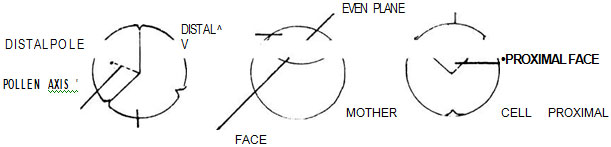 Fig. 12.2 A. Polar axis; B. Proximal pole and distal pole; C. Even plane
Fig. 12.2 A. Polar axis; B. Proximal pole and distal pole; C. Even planePolar spores are isopolar, subisopolar or heteropolar. An even plane is a line considered to be situated halfway between the poles and cutting the polar axis at right angles (Fig. 12.2 C). In isopolar spores such an even plane divides the spores into equal halves, the proximal and the distal half. The surface of the proximal half is known as the proximal face, and the surface of the distal half as the distal face. Hence, in isopolar spores, the two faces are similar with regard to apertures etc. In heteropolar spores, the proximal and distal faces are pronouncedly dissimilar to each other with regard to apertures etc. Subisopolar spores are somewhat intermediate between isopolar and heteropolar. Their equatorial plane is usually more or less curved.
Spores are asymmetric or symmetric.
Symmetric spores are either radiosymmetric or bilateral, and are of common occurrence. The radiosymmetric spores have more than two vertical planes of symmetry, i.e., they can be divided into two equal halves along any vertical plane. If a radiosymmetric spore be provided with only two such vertical planes, it has always equilong equatorial axes. Bilateral spores have two vertical planes of symmetry; however, as compared with the radiosymmetric spores, the equatorial axes are not equilong, e.g., Cryptomeria, Isoglossa.
The asymmetric spores are of rare occurrence. They are either non-fixiform (without fixed shape) or fixiform (with -fixed shape).
Erdtman (1945) suggested size classes based on the length of the longest spore axis while considering the size of the spore; spines and other excrescences rising beyond the general surface of the spores are usually not included. It is sometimes difficult to define the "general surface" of the spore. Hence, in such cases, the methods used in obtaining the measurements are stated. It may sometimes be profitable to quote the dimensions of the spore lumen. The size classes suggested by Erdtman are as follows:
Very small spores (sporae perminutae - PI) < 10 m
Small spores (sporae minutae - MI) 10-25 m
Medium size spores (sporae mediae - ME) 25-50m
Large spores (sporae magnae - MA) 50-100 m
Very large spores (sporae permagnae - PA) 100-200 m
Gigantic spores (sporae gigantae - GI)> 200 m
The shape of radiosymmetric spores, that are equatorially unconstricted, is ellipsoidal, and in equatorial view, the outline is circular or somewhat elliptical. With the polar axis comprising the major axis of the ellipse, the shape of the pollen grains with increasing eccentricity may be spherical, prolate spheroidal, subprolate, prolate and perprolate. In case of those pollen grains in which the polar axis comprises the minor axis of the ellipse, the shape of the pollen grains with increasing eccentricity may be spherical, oblate spheroidal, suboblate, oblate and peroblate. If P represents the polar axis of the pollen grain and E represents the other axis of the ellipse, i.e., the line marking the total breadth of the grain in equatorial view then the P/E ratio, i.e., relation between P and E helps to decide the shape classes. The suboblate, oblatespheroidal, prolate spheroidal and subprolate pollen grains may be grouped together as sub-spheroidal. Accordingly, the main shape classes can be peroblate, oblate subspheroidal, prolate and perprolate with respective P/E ratios as < 4/8, 4/8-6/8, 6/8-8/6, 8/6-8/4 and > 8/4.
Aperture is an opening or thin, more or less distinctly delimited area which is normally engaged in the discharge of soft inner spore material or part of it. In other words, aperture of a spore is any weak, preformed part of the general surface of the spore which may be engaged in forming an opening in connection with the normal exit of intra-exinous substance or for coming out of the pollen tube. Spores are usually provided with apertures, i.e., they are aperturate; they are rarely non-aperturate. In classification of apertures, their position, shape, structure, number and size have to be considered.
Position and Shape of apertures
The apertures can be grouped into three types - polar, global and equatorial.
Polar apertures: Polar apertures can be monopolar (proximalipolar or distalipolar) or bipolar. The spores of mosses and ferns with tetrad scars (laesurae) are examples of proximalipolar apertures. Spores are monolete if they have one laesura and trilete if they have three laesurae radiating from the proximal pole.
Sulcoid apertures which neither reach nor pass across the distal pore is termed as sulculi (singular sulcus - small sulcus). They are situated in the distal face of the grains (sometimes even near the equator). A bi-sulculate pollen grain has two sulcoid furrows, which are parallel to each other, and with the distal pole in the centre, (between the furrows). In trisulculate grains, the sulcute stand in relation to the pole in same way as the sides of an equilateral triangle to the centre of the triangle.
Equatorial or subequatorial apertures are of two types - pori and colpi. A porus is a circular aperture placed along the equatorial region. There may be only one porus. The spore is then called monoporate (Fig. 12.3).
If there is a meridional, elongate aperture with one of its axes (usually the congest) crossing the equator at right angles, then it is called colpa A typical monocotyledonous pollen is monocolpate, and a dicotyledonous pollen is tricolpate. The rest of the non-aperturate surface of a colpate grain is called apocolpia. Similarly, in a porate grain, the non-aperturate surface is called apoporia.
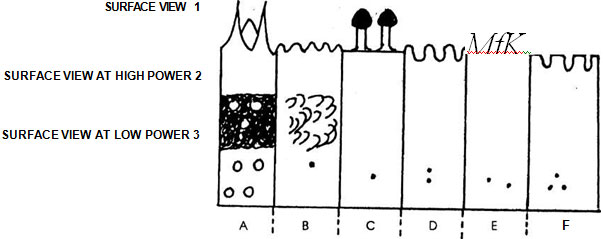 Fig. 12.3 Shape and position of aperture
Fig. 12.3 Shape and position of apertureA. Echinate; B. Granular; C. Piliferous; D. Scrobiculate (pitted); E. Reticulate; F. Negative reticulum (exine isles separated by network of grooves)
The pollen grains of Berberis, Eriocaulon are provided with spiral or spiruloid apertures or with somewhat aperturoid weak streaks which are probably derived from colpi, rugae, or sulci etc. Such grains are called spiraperturate grains.
In certain aperture types, the successive openings are not congruent. The superficial parts of the apertures have the shape of colpi, rugae or pori, whereas the underlying parts are either circular, longitudinally elongate (lolongate) or transversely elongate (lalongate). Thus they exhibit the shape of a mouth and are termed ora. Grains with lalongate ora meeting laterally to form a continuous oral zone are called zonorate. Grains with composite apertures are thus named after the corresponding simple aperture types and referred to as colporate, rugorate, pororate, etc.
The apertures are usually covered with aperture membranes. When the membrane is smooth, it is termed psilate, when it is provided with granules, granulate and when it is thickly beset with coarse granules, then called as crustate.
The newly developed microspore consists of centrally situated nucleus, dense cytoplasm, and the spore wall (sporoderm). The sporoderm usually consists of two strata - the inner, soft layer is the intine, and the outer, hard one the exine or sclerina. Though sclerine is synonymous with exine, in certain mosses and ferns, there is also another layer called pezine.
The exine appears at first as a delicate new membrane, but later on it thickens considerably and becomes differentiated into two layers - the outer sexine (equivalent to perine of kerner, and also sometimes called exo-extine), and the inner, nexine (endo-extine). Sexine is the outer, sculptured part of the exine. It is further differentiated into two layers, viz., the inner endosexine and the outer ectosexine. The inner, non-sculptured part of the exine is the nexine which is further differentiated into ecotonexine and endo-nexine. The nexine is thick, uniform, and continuous except below the pores.
According to Erdtman, the external surface of the pollen may be characterised by various markings such as pilate, reticulate, striate, ornate, crassis-exinous, tenui-sexinous, subsacatte, tegillate (verrucose and spinose) etc., and are referred to as exine ornamentations or sexine patterns.
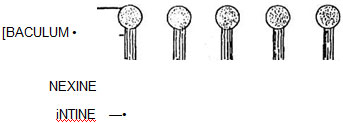 Fig. 12.4 Basic elements of sexine
Fig. 12.4 Basic elements of sexineSexine patterns
The different types of ornamentation of exine is defined as sexine patterns. Among flowering plants one of the basic elements of ^ -I the sexine pattern appears to consists of small K drum-stick shaped rods called pila (sing. pilum) projecting at right angles from the nexine surface. Each pilum has a head called caput which is supported by a rod-like neck called baculum (Fig. 12.4). The capita (pi. of caput) form the upper part of the sexine, i.e., ectosexine and bacula form the basal part, i.e., endosexine. If the capita amalgamate (unite), a layer will be formed on the top of the pila joining the capita and leaving the bacula (tegillum - small roof) free (fig. 12.5). A tegillate sexine is of different types. If the ectosexine is atleast twice as thick as the endosexine, then the tegillate sexine is crossitegillate (Fig. 12.6); if it is atleast twice as thin as, i.e., half the endosexine, then tegillate sexine is tenuitegillate (Fig. 12.7). Tegillate sexine is referred to as punctitegillate if it is provided with small, perpendicular holes (puncta - Fig. 12.8).
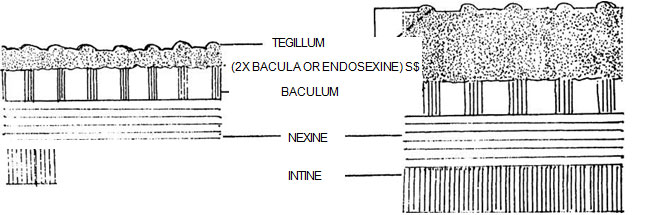 Fig. 12.5 Formation of tegillum
Fig. 12.5 Formation of tegillumFig. 12.6 Crossitegillate sexine
Punctitegillate sexine is found in the ollen grains of many plants like Opuntia. The upper surface of a tegillum is usually psilate (smooth) or slightly undulating (wavy) in the grains with unditegillate sexine (Fig. 12.9). It is sometimes beset with sculptered elements in shape of granules (granula), warts (verrucae), spines (spinae), spinules (spinulae). They are also described as echinate (spiny) subechinate (spinules), verrucate (warts), granulate (granules), piliferous (pila), scrobiculate (pits) striate (striate) and reticulate (reticulation).
In saccate pollen grains, i.e., grains provided with air sacs (saccri) and in subsaccate grains (air sacs in the form of small bladders or irregular waves and crests) the bacula loose locally their foothold in the nexine.
A striate sexine pattern is formed by a lateral union of pila (or their capita) standing in a rows.
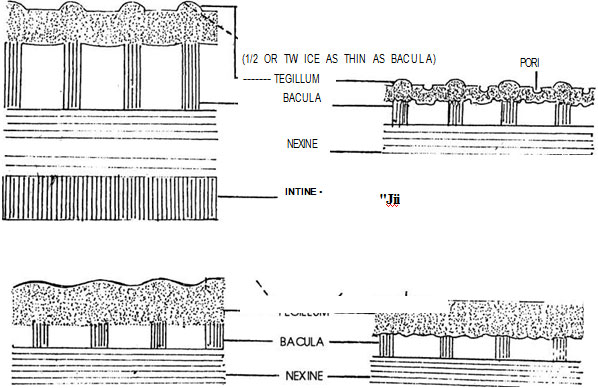 Fig. 12.7 Tenuitegillate sexine
Fig. 12.7 Tenuitegillate sexineFig. 12.8 Punctitegillate sexine
 Fig. 12.9 Unditegillate sexine
Fig. 12.9 Unditegillate sexineFig. 12.10 Psilate sexine
A reticulate sexine pattern consists of meshes (brochi).
Certain grains, though not common are oligobrachate, i.e., provided with a few meshes while others are mesopolybrochate, i.e., provided with large number of meshes. The reticulum may be of one size and known as homobrochate or of different sizes called heterobrochate.
A reticulum is also said to consist of walls, the muri and spaces, the lumina between the walls. According to the number and arrangement of bacula in the muri it is possible to distinguish between simplibaculate, duphibaculate and multibaculate muri; i.e., bacula are arranged in one, two or many rows. If the muri are not supported by individual bacula but by bacularia (groups of fused bacula), they may be termed as simpli, dupli, multibaculariate according to the arrangement of bacularia. In an angustimurate reticulum, the thickness of the muri is one fifth of the diameter of the lumina or lesser; in a latimurate reticulum, it is as thick as the diameter of the lumina or thicker.
Muri are not always continuous. If, as in Cuscuta and Liliaceae plants, they are composed of isolate pila in polygonal arrangement, the pattern is called retipilate. In ornate grains, muri are curved and occasionally interrupted forming different patterns.
Pollengrains are said to be areolate if the sexine is chiefly or exclusively confined to areas (areolae) which correspond in size and position to the lumina of the reticulate sexine.
Excrescences
In some cases the exine bears outgrowths of various types on its roof or tegillum. As regards the origin and growth, such outgrowths may or may not be real outgrowths from the exine, and are further called as excrescences. These outgrowths constitute the sculptural elements. Hence, sometimes the ectosexinets forming the tegillum (roof) by amalgamation of the capita of the pila is beset with excrescences. These may be in the form of small granules called granula and the exine is termed granulose, in the form of warts (verrucae) exine being called verrucose; in the form of large and conspicuous spines (spinae), spinose; sometimes excrescences are in the form of small, short spines, the spinules (spinulae), forming spinulose exine.
Palynogram
A palynogram is a diagrammatic representation of a pollen grain or microspore, regarding its symmetry apertures, shape, size, sporoderm stratification, sexine patterns, etc. Thus, the chief results of morphological analysis of a pollen grain or a spore can be exhibited by a palynogram. In a palynogram, the spores with distinct polarity are shown either in equatorial or in polar view.
In case of a palynogram, drawn in equatorial view of an isopolar, radiosymmetric pollengrain, with equatorial apertures, the centre is occupied by an aperture. In polar view of the same spore, one of the poles is exactly at the centre of the palynogram with one of the apertures vertically below it.
In a palynogram the surface of sexine is shown as an optical section passing through the spore-wall is also exhibited.
Palynology in relation of Honey Industry (Melittopalynology)
Honey was man's first sweetening material. Honey contains 70 to 75% invert sugars, together with proteins, minerals, salts and water. Most showing flowers secrete nectar, which serves to attract insects for pollination. Nectar is composed chiefly of sucrose, with some fructose and glucose. After partial digestion, nectar is converted into honey, and in this form, it is stored for future use.
Honey is used as food by bees. Honey bees derive their food in the form of nectar and pollens. While collecting the nectar and pollens they pollinate the plants. Pollination by honey bees is a good example of symbiosis. It is reciprocal, nutritive, disjunctive symbiosis wherein the honey bee obtains food in the form of nectar and pollen from the flowers while pollinating them. The honey bees are in certain respects the most efficient. The honey bees are in certain respects the most efficient of all pollinating insects. This is partly due to the fact that nectar is secreted in nectaries, and bees select those plants which have the maximum sugar contents (70% to 80%) in the nectar. The pollen supplies food in the form of proteins, fats, carbohydrates, minerals, vitamins, enzymes, hormones etc. According to the source of nectar, there is variation in the flavour and quality of honey. Bees fix their preference for a particular plant species and thus show floral fidelity. Although many plants are visited by bees, some are especially favoured, and these are, afterwards cultivated near apairies; e.g., Pista, Jamb and Kavi as used at Mahabaleshwar. From the samples of honey studied, the pollens of Eugenia, Putragiva, Citrus, Plectranthus, Brassica, Sapindus etc., they work continuously from the beginning to the end of the flowering season, partly due to the fact that they visit flowers for both pollen and nectar and partly because of their remarkable precision which leads them to visit a large numbers of flowers of the same species in rapid succession. A honeybee, for example, visits only one kind of flower on any collecting trip and usually on any one day. Furthermore, the bees have hairy legs that are well fitted for carrying pollen and their long, tongue-like probosces which enable them to obtain nectar from irregular, tubular and partly closed corollas are recorded. The anemphilous plants like Holoptelia, Borassus and Cocos also provide pollen food to the young larvae of insects.
Besides the pollengrains, the honey sample may also contain fungal spores. This may be due to
- deliberate collection by bees to substitute pollens,
- incidental collection due to the collection of pollens contaminated by fungal spores or
- wind-borne fungal spores.
Sometimes, honey gets contaminated with poisonous pollens; e.g., pollen of Eugenia geniculata. Such honey may be toxic to both the bees as well as the humans or it may be toxic only to human.
Diagnosis of honeys
Diagnosis of honey is done to check bee-diseases caused by pollen toxicity, contamination due to fungal spores, or to detect poisonous honey, i.e., poisonous to human beings. Honey diagnosis protects the consumers.
The analysis of honey samples reveals the type of honey and its geographical origin.
Types of honey
When a honey sample contains pollengrains of only a single genus or species of flowering plants, then it is called unifloral honey. Such honey is usually marketed under the name of the plant source, e.g., 'Jambul' honey, 'Gela' honey etc. When the honey sample contains pollengrains belonging to different plant sources it is called multi-floral honey.
Geographical origin of honey
There are a considerable number of plants that can be pollinated only by bees and such plants are limited in their distribution to those regions that are inhabited by bees. A spectrum of pollen types represented in a honey samples of local honey often reveals the local flora which has a characteristic plant association. This often enables to identity the geographical origin of the honey sample. The honey is then marketed sometimes under the title of the locality, e.g., Kashmir honey, Nepal honey, Mahabaleshwar honey, etc.
Support our developers




