Propagation of Metabolic Perturbations through Networks
The metabolic consequences of altering the amount of an enzyme are unlikely to
be confined to a single pathway. A clear illustration of the extent of the interactions
that occur between pathways is provided by a study of transgenic tobacco
lines in which the amount of transketolase was selectively decreased (Henkes
et al., 2001). These lines displayed a near-proportional decrease in the maximum
rate of photosynthesis in saturating CO
2 and a smaller inhibition of photosynthesis
under normal growth conditions. This inhibition was accompanied by large
decreases in the steady-state levels of RuBP and 3PGA, smaller decreases in the
amounts of triose phosphates and fructose 1,6-bisphosphate, and a large increase in
the amount of fructose 6-phosphate. These changes are entirely consistent with
restrictions in the two reactions of the Calvin cycle catalyzed by transketolase and
suggest that the immediate cause for the decrease in photosynthesis is a restriction
in the ability to regenerate RuBP (Fig. 1.3).
Thus, the effect of reduced transketolase
appears to be similar to that obtained when the aldolase content was decreased
under low light (Fig. 1.2A).
However, in contrast to the consequences of manipulating
aldolase content, a decrease in transketolase also caused a disproportionately
large decrease in the levels of aromatic amino acids, intermediates of the
phenylpropanoid pathway, and secondary products such as chlorogenic acid and
lignin. These observations suggest that the level of transketolase has a major impact on the channeling of intermediates into the shikimic acid pathway and the likely
explanation for this effect is that the metabolic network responds to a decrease in
the amount of transketolase by decreasing the amount of erythrose 4-phosphate
(Fig. 1.3). Consequently, flux into the shikimic acid pathway is restricted by
the supply of erythrose 4-phosphate and phenylpropanoid metabolism is
constrained by the corresponding decreased provision of aromatic amino acids.
The multiple responses to reducing transketolase highlight the extent of integration
within the central metabolic pathways and the potential difficulties
in attempting to modify flux through a specific section of the metabolic network.
In particular, the results suggest that major changes in secondary metabolism may
require appropriate reprograming of primary pathways to ensure an adequate
supply of the necessary precursors. Corroborative evidence that the formation of
secondary products may be limited by the availability of primary precursors is
provided by a report that a decrease in the levels of aromatic amino acids due to
ectopic expression of tryptophan decarboxylase led to decreases in the amounts of
chlorogenic acid and lignin in transgenic potato plants (Yao
et al., 1995).
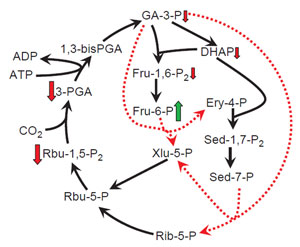 |
| FIGURE 1.3 Effect of decreased transketolase
content on photosynthetic intermediates in
tobacco plants (Henkes et al., 2001). Changes in
the steady-state levels of Calvin cycle
intermediates
in transketolase-antisense lines are
compared with those in wild-type plants grown
under the same conditions. The reactions
catalyzed by transketolase are indicated by
dotted lines.
Symbols refer to the following
changes inmetabolite content: ↑, increase; ↓,
decrease. |
In fact both the structure and chemical organization of metabolic networks
suggest that transketolase is unlikely to be unique in the manner in which changes
in its activity influence other metabolic processes. This view is supported by a
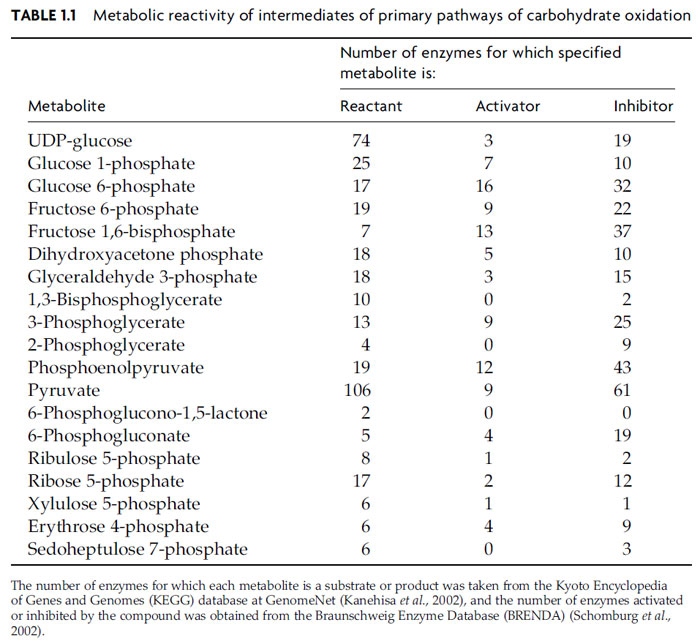
theoretical analysis of the potential metabolic interactions for each of the intermediates
of glycolysis and the oxidative pentose phosphate pathway (Table 1.1).
Although there is considerable variation between compounds, on average each
metabolite is a reactant for about 20 enzymes, and either activates or inhibits a
further 22 enzymes. These values provide only a crude estimate of the complexity
that arises through the multiplicity of ligand-binding interactions and the estimate is in any case very dependent on the extent to which all potential inhibitory and
stimulatory responses have been identified for the selected enzymes. Even so, the
analysis suggests that perturbation of the level of any metabolite within the central
pathways of carbohydrate oxidation has a very strong likelihood of affecting
several other reactions, thus allowing the consequences of the initial change to
propagate widely throughout the network. Such considerations further emphasize
the integrated nature of the central metabolic pathways and the difficulties that are
likely to be encountered in attempting to modify individual processes selectively.
 |





