The Lacertilia
Some few Lacertilia, like the Chamaeleons and the Amphisbaence, are covered by a soft integument; but, in the majority, there is an epidermic exoskeleton composed of homy plates, tubercles, or spines, or overlapping scales. In some forms (e. g., Scincus, Cyclodus) the dermi beneath the horny scales is ossified, and the body has a complete armor of bony scutes, corresponding in form with the scales. The dermal ossifications of the head may coalesce with the subjacent bones, but this union of dermal bones with subjacent parts does not occur in other parts of the body.The spinal column always contains a considerable number of vertebrae; and, except in the Amphisbaene and some few other Lizards, the tail is long. Those Lizards which possess hind-limbs have a sacrum, into which not more than three vertebrae, and rarely more than two, enter. The presacral vertebrae are distinguishable, when sternal ribs are present, into cervical and dorsal. All those vertebrae which lie in front of the first sternal rib are cervical; and if, as sometimes happens, the last two or three dorsal vertebrae are devoid of ribs, they become lumbar. Not more than nine vertebrae are met with in the cervical region of existing Lacertilia, and this number is rare. The number was greater in some extinct Lacertilia.
The atlas is composed of three pieces, one inferior and two superolateral. The odontoid bone is closely united with the second vertebra, and its anterior face may be cylindroidal. A separate ossification is sometimes formed on the under-surface of the spinal column at the junction of each pair of vertebrae. Such a separate ossification, or sub-vertebral wedgebone, is commonly developed beneath and between the odontoid bone and the body of the second vertebra.
The centra of the vertebrae are either procoelous, or amphicoelous; the former being by far the more common condition in existing Lacertilia, all of which, except the Geckos and Sphenodon, have procoelous vertebrae. The cups and balls are usually ellipsoidal, the long axis of the ellipsoid being transverse. In the Geckos, the centra of the vertebrae are conically excavated at each end; and, except in the centre of each vertebra, where it is ossified, the notochord persists throughout the spinal column.
The sacral vertebrae of existing Lacertilia are not anchylosed together, nor are their articular faces modified, the two being connected by a free cup-and-ball articulation. The movements of the two vertebrae, however, are restrained by the strong ligaments which connect their neural spines and arches, and by the fibro-cartilage which connects and covers the free ends of their expanded ribs.
In the anterior part of the tail of the Lacertilia there are usually well-developed subvertebral chevron bones, which are commonly attached to the bodies of the several vertebrae, and not in the intervals between adjacent vertebrae. In many Lacertilia (Lacertae, Iguanae, Geckos) the caudal vertebrae have a very singular structure, the middle of each being traversed by a thin, unossified, transverse septum. The vertebra naturally breaks with great readiness through the plane of the septum, and when such Lizards are seized by the tail, that appendage is pretty certain to part at one of these weak points.
The arches of the vertebrae of the Lacertilia are articulated together by the ordinary oblique processes, or zygapophyses. In the Iguanae. they are additionally connected by a process of the front part of each arch (zygosphene), which fits into a fossa on the posterior face of the preceding arch (zygantrum). These Lacertian vertebrae thus nearly approach the vertebrae of the Ophidia.
The transverse processes of the vertebrae are very short, and are, at most, divided into two indistinct facets, with which corresponding facets on the proximal ends of the ribs articulate.
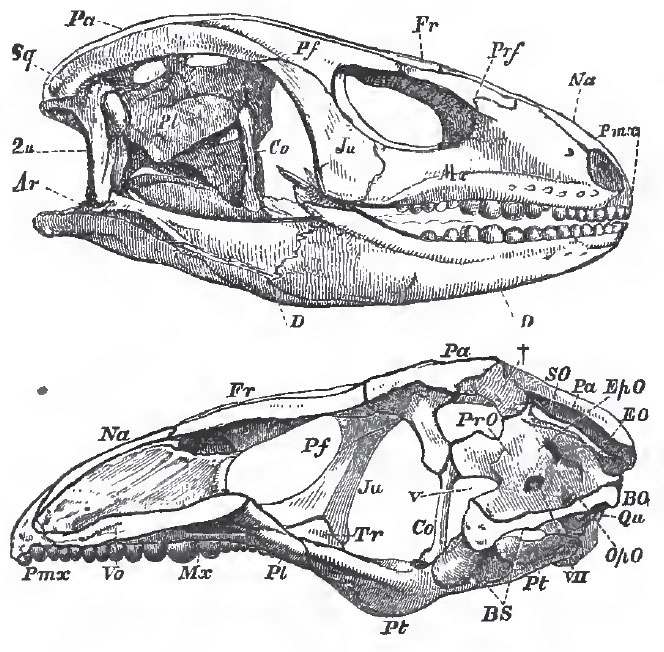 |
| Fig. 69. - The skull of Cyclodus, entire and longitudinally bisected. |
The Flying Lizard (Draco volans) is remarkable for the elongation of many of its posterior ribs, which are continued into, and support, the parachute-like expression of the integument by which it is enabled to perform its flights.
The skull of the Lacertilia resembles that of the Chelonia in the development of an interorbital septum (except in the Amphisbaenae), and in the absence of alisphenoids, or of any complete ossification of the presphenoidal or orbito-sphenoidal regions. The premaxillae and maxillae are firmly united with one another and with the skull, and there are two vomers. An unossified space, the parietal foramen, usually remains in the roof of the skull in the course of the sagittal suture, or between the parietals and the frontals.
In the principal group of the Lacertilia, a column-like membrane-bone, called the columella (but which is not to be, by any means, confounded with the stapes, to which the same name is often applied in Reptiles), extends from the parietal to the pterygoid on each side, in close contact with the membranous or cartilaginous wall of the skull. Hence they have been called "Kionocrania," or "column skulls." This columella (Fig. 69, Co) appears to correspond with a small independent ossification, which is connected with the descending process of the parietal and with the pterygoid, in some Chelonia.
In the great majority of the Lacertilia (as in the Chelonia), the side-walls of the skull, in the region of the ear, are produced into two broad and long parotic processes, into the composition of which the opisthotic, ex-occipital, and prootic bones enter. Each quadrate bone is articulated with the outer end of one of these processes (in which a small separate pterotic ossification sometimes appears), and is usually movable. The parietal bones do not unite suturally with the occipital segment of the skull, or with the prootic bones, but are connected with them only by fibrous tissue. And as the presphenoidal region remains unossified, or incompletely ossified, it follows that the fronto-parietal portion of the skull is, in most Lizards, slightly movable upon the occipito-sphenoidal part.
Each parietal bone is prolonged backward into a process which articulates with the upper part of the parotic prolongation of the skull; and to the outer side of the posterior extremity of the parietal process the squamosal is attached. The squamosal may be continued forward to the post-frontal, which is sometimes subdivided into two. The post-frontal may unite below with the jugal, and thus bound the orbit. Only in Sphenodon, among recant Lizards, is the jugal connected with the distal end of the quadrate by bone. As a general rule, the quadrato-jugal is represented only by a ligament.
In consequence of the structure which has been described, the posterior region of the ordinary Lacertilian skull presents a number of distinct fossae in the dry state. A supra-temporal fossa lies between the parietal, the post-frontal, and the squamosal, on the upper face of the skull; a post-temporal, between the parietal, the occipital, and the parotic apophysis on the posterior face; a lateral-temporal, between the squamosal and post-frontal above, the jugal and quadrate in front and behind, and the quadrato-jugal ligament below.
The palatine and pterygoid bones are firmly connected both with the facial bones, and with the floor of the skull. Thus the basisphenoid gives off two basipterygoid processes, the outer ends of which articulate with the inner sides of the pterygoid. The posterior ends of the pterygoids are usually connected with the inner surfaces of the distal ends of the quadrate bones. Their anterior ends are firmly united with the palatines; and, from the junction of the two, a transverse bone (Fig. 70, Tr) usually passes, to unite the palatine and pterygoid with the maxilla.
The anterior ends of the palatines unite with the maxillae and the vomers; but, in existing Lacertilia, they do not meet one another, or come into contact with the basisphenoid or presphenoid in the middle line. The palatine apertures of the nostrils are placed between the palatine bones, on the outer side, and the vomer, on the inner. In only a few Lacertilia do the palatine bones send down processes which bend toward one another in the middle line, and so form a posterioi nasal passage, partially separated from the oral cavity.
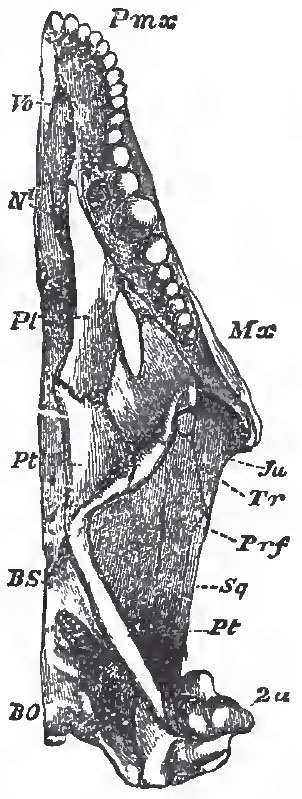 |
| Fig. 70. - Under-view of the skull of Cyclodus: N posterior nasal aperture |
The hyoidean apparatus consists of an elongated median rod, the anterior part of which supports the base of the tongue; and, usually, of two long cornua on each side of this. The cephalic ends of the anterior cornua may be perfectly free, and lie upon the sides of the neck, as in Psammosaurus; or they may be traceable to, and be connected with, the stapes and the parotic processes, as in Sphenodon.
The limbs may be completely developed; or only one pair (either the anterior or the posterior) may be present; or they may be entirely absent. When present, they may be mere styliform rudiments, or may possess any number of digits from two to five. Even when the limbs are altogether absent, the pectoral arch remains, though the pelvic arch seems to vanish. When the pectoral arch is complete, it consists of a suprascapula, scapula, coracoid (with precoracoid and epicoracoid elements), and two clavicles, united by an interclavicle, which lies in a groove of the sternum. (Figs. 12 and 13, pp. 35 and 36.)
The coracoids articulate with grooves in the anterolateral edges of the sternum, and usually more or less cross and overlap one another, in front.
In the genus Lialis, in which not a trace of a fore-limb is discernible, there is a small sternum, consisting of a flat, somewhat pentagonal, plate of cartilage, in which there is a little coarsely-granular calcareous deposit; but this sternum is connected with no ribs, nor, though it lies between the coracoids, does it articulate with them. Each coraco-scapular arch is a continuous cartilage, narrow in the middle, but expanded at its dorsal, and still more at its sternal end, where the right overlaps the left, and both are connected by fibrous tissue with the sternum. The narrow middle part of the coracoid is invested, and in part replaced, by a sheath of membrane bone, which expands above and below, and represents both scapula and coracoid, though it presents no trace either of division, or of a glenoidal cavity. Beyond the extremities of this central ossification the cartilage merely presents scattered granular calcification. Along the front edge of each coracoscapular arch, and closely connected with its ossified part, is a long curved clavicle, entirely composed of membrane-bone, and united with its fellow in the ventral median line, by ligamentous fibres. There is no interclavicle. The pectoral arch in other snake-like Lizards, such as the Blind-Worm (Anguis) and the Sheltopusik (Pseudopus), is in much the same condition as in Lialis.
When the hind-limbs are well developed, there is a complete pelvis. The ilia are movably articulated with the fibrecartilages which cover the ends of the sacral ribs. The pubes and the ischia meet in median symphyses, and the anterior margin of the pubis usually, as in the Chelonia, gives off a strong curved process. In many Lacertilia a partially-ossified or cartilaginous rod (os cloacoe) is continued back from the symphysis of the ischia, and supports the front wall of the cloaca.
In most Lacertilia the manus possesses five digits; and, when this is the case, there are usually eight bones in the carpus-one for each metacarpal on the distal side, one radial, one ulnar, and one central. As a very general rule, the pollex has two phalanges, the second digit three, the third four, the fourth five, and the fifth three (2, 3, 4, 5, 3). The pes, also, generally possesses five digits, which increase in length to the fourth, the fifth being smaller than the rest, and divergent in direction. Two large bones, very closely united, or completely fixed together, represent the calcaneum and the astragalus, and are articulated, in a manner which allows of very little motion, with the tibia and fibula. In the distal row there is usually a large bone, representing the cuboid. The fifth metatarsal (The bone thus named may perhaps contain a tarsal element, and represent not only the fifth metatarsal, but the corresponding distal tarsale.) is bent, as in the Chelonia, and may articulate with the calcaneum as well as with the cuboid. One or two of the cuneiform bones may be present, or the inner ones may be represented merely by fibrous membrane, or by cartilage; in which latter case the inner metatarsals appear to articulate directly with the astragalus in the skeleton. The number of the phalanges is very generally the same as in the manus for the four tibial toes, but one more for the fibular (2, 3, 4, 5, 4).
The Lacertilia all possess teeth, which may be confined to the premaxillae, maxillae, and dentary piece of the mandibles; or may, in addition, be developed on the palatine and pterygoid bones. These teeth are simple in structure, and their crowns have very various forms, being sometimes sharp and conical (Monitor); or blade-like, with serrated edges (Iguana); or with broad, crushing, and spheroidal crowns (Cyclodus). As a general rule, the teeth become anohylosed to the adjacent bone with age; and in the upper and lower jaws they thus become attached, either by their sides to the parapet of the jaw, when the dentition is said to be pleurodont; or by their bases to the summit of the parapet, when the dentition is acrodont. The extinct Protorosauria are said to be thecodont, or to have the teeth lodged in alveoli. New teeth are usually developed at the bases of the old ones.
The Lacertilia are divisible into numerous groups, the leading distinctive characters of which are exhibited in the following table:
I. - The pterygoid and quadrate bones united.
- A columella and an interorbital septum in the skull. Kionocrania (Stannius).
- Amphicoelous vertebrae (K. Amphicoelia).
- Dentition acrodont or pleurodont.
1. Ascalabota.
2. Rhynchocephala.
3. Homoeosauria (The columella has not been observed in these groups.) - Dentition thecodont (?) 4. Protorosauria. (The columella has not been observed in these groups.)
- Procoelous vertebrae (K. procoelia).
- Not more than nine cervical vertebrae.
- The nasal bone, single. 5. Platynota.
- The nasal bones, two.
- The integument of the head not covered with epidermic plates. 6. Eunota.
- The integument of the head covered with epidermic plates. 7. Lacertina.
- More than nine cervical vertebrae. 10. Dolichosauria. (The columella has not been observed in these groups.)
- No columella; no interorbital septum. 12. Amphisbaenoida.
8. Chalcidea.
9. Scincoidea.
11. Mosasauria.
II. - The pterygoid and quadrate bones united.
13. Chamaeleonida.Support our developers




