Gene Regulation
The DNA of a microbial cell consists of genes a few to thousands which do not express at the same time. At a particular time only a few genes express and synthesize the desired protein. The other genes remain silent at this moment and express when required. Requirement of gene expression is governed by the environment in which they grow. This shows that the genes have a property to switch on and switch off.
Twenty different amino acids constitute different protein. All are synthesized by codons. Therefore, synthesis of all the amino acids requires energy which is useless because all the amino acids constituting proteins are not needed at a time. Hence, there is need to control the synthesis of those amino acids (proteins) which are not required. By doing this the energy of a living cell is conserved and cells become more competent. Therefore, a control system is operative which is known as
gene regulation.
There are certain substrates called inducers that induce the enzyme synthesis. For example, if yeast cells are grown in medium containing lactose, an enzyme lactase is formed. Lactase hydrolyses the lactose into glucose and galactose. In the absence of lactase, lactase synthesis does not occur. This shows that lactose induces the enzyme lactase. Therefore, lactase is known as inducible enzyme. In addition, sometimes the end product of metabolism has inhibitory effect on the synthesis of enzyme. This phenomenon is called
feedback or
end product inhibition.
From the outgoing discussion it appears that a cell has auto control mediated by the gene itself. For the first time Francois Jacob and Jacques Monod (1961) at the Pasteur Institute (Paris) put forward a hypothesis to explain the induction and repression of enzyme, synthesis. They investigated the regulation of activities of genes which control lactase fermentation in
E. coli through synthesis of an enzyme, P-galactosidase. For this significant contribution in the field of biochemistry they were awarded Nobel Prize in Medicine in 1965.
Gene expression of prokaryotes is controlled basically at two levels
i.e. transcription and translation stages. In addition, mRNA degradation and protein modification also play a role in regulation. Most of the prokaryotic genes that are regulated are controlled at transcriptional stage. Other control measures operating at different levels are given in Table.2.4.
Transcriptional Control
It is a general strategy in a living organism that chemical changes occur by a metabolic pathway through a chain of reactions. Each step is determined by the enzyme.
Again synthesis of an enzyme comes under the control of genetic material
i.e. DNA in living organisms. Enzyme (protein) synthesis occurs via two steps : transcription and translation. Transcription refers to synthesis of mRNA. Transcription is regulated at or around promoter region of a gene. By controlling the ability of RNA polymerase to the promoter the cell can modulate the amount of message being transcribed through the structural gene. However, if RNA polymerase has bound, it again can modulate transcription. By doing so the amount of gene product synthesized is also modulated. The coding region is also called structural gene. Adjacted to it are regulatory regions that control the structural gene. The regulatory regions are composed of promoter (for the initiation of transcription) and an operator (where a diffusible regulatory protein binds) regions.
The molecular mechanisms for each of regulatory patterns vary widely but usually fall in one of two major groups: negative regulation and positive regulation. In negative regulation an inhibitor is present in the cell and prevents transcription. This inhibitor is called as repressor. An inducer
i.e. antagonist repressor is required to permit the initiation of transcription. In a positive regulated system an effector molecule (
i.e. a protein, molecule or molecular complex) activates a promoter. The repressor proteins produce negative control, whereas the activator proteins produce positive control. Since the transcription process is accomplished in three steps polymerase binding, isomerization of a few nucleotides and release of RNA polymerase from promoter region), the negative regulators usually block the binding, whereas the activators interact with RNA polymerase making one or more steps. In negative regulation an inhibitor is bound to the DNA molecule. It must be removed for efficient transcription. In positive regulation an effector molecule must bind to DNA for transcription.
Table 2.4. Types of gene regulation in prokaryotes operating at different levels.
|
Levels of Control |
|
Means of Control |
|
Examples |
| 1. |
Transcriptional control |
1. |
DNA binding proteins |
(i) |
Helix-turn-helix in phage λ |
| |
|
|
|
(ii) |
Zinc fingers in Xenopus |
| |
|
2. |
The lac operon |
|
E.coli |
| |
|
3. |
Catabolic control |
|
Cyclic AMP in E.coli |
| |
|
4. |
The gal operon |
|
|
| |
|
5. |
The arabinose operon |
|
E.coli |
| |
|
6. |
The trp operon |
|
E.coli |
| 2. |
Translation control |
|
The arg regulon |
|
E.coli |
| 3. |
The membrane- mediated
regulation |
|
The put system |
|
Salmonella typhimurium |
| 4. |
Osmotic control |
|
Turgor |
|
E.coli, etc. |
| 5. |
Through Electron transport |
|
Stringent control |
|
Response to amino acid starvation |
The Lac Operon
For the first time Jacob and Monod (1961) gave the concept of operon mode! to explain the regulation of gene action. An operon is defined as several distinct genes situated in tadem all controlled by a common regulatory region) Commonly an operon consists of repressor, promoter, operator and structural genes. The message produced by an operon is polycistronic because the information of all the structural genes resides on a single molecule of mRNA.
The regulatory mechanism of operon responsible for utilization of lactose as a carbon source is called the
lac operon.lt was extensively studied for the first time by Jacob and Monod (1961). Lactose is a disaccharide which is composed of glucose and galactose.
The lactose utilizing system consists of two types of components; the structural genes
(lacZ, LacY and
lac A) the products of which are required for transport and metabolism of lactose and the regulatory genes (the
lacI
, the
lacO and the
lacP
). These two components together comprises the
lac operon (Fig. 2.11). One of the most key features is that operon provides a mechanism for the coordinate expression of structural genes controlled by regulatory genes. Secondly, operon shows polarity
i.e. the genes Z,Y and A synthesize equal quantities of three enzymes b-galactosidase (by
lacZ), permease ( by
lacY) and acetylase (by
lacA). These are synthesized in an order
i.e. b-galactosidase first and acetylase in the last.
(i) The structural genes. The structural genes form one long polycistronic mRNA molecule. The number of structural gene corresponds to the number of proteins. Each structural gene is controlled independently, and transcribe mRNA molecules separately. This depends on substrates to be utilized. For example, in
lac operon three structural genes (Z,Y and A) are associated with lactose utilization (Fig. 2.11). b-galactose is the product of
lacZ that cleaves b-1,4 linkage of lactose and releases the free monosaccharides. This enzyme is a tetramer of four identical subunits each with molecular weight of 1,16,400. The enzyme permease (a product of
lacY) facilitates the lactose to enter inside the bacterium. Permease has molecular weight of 46,500. It is hydrophobia The cells mutant in
lacZ and
lacY are designated as Lac"
i.e. the bacteria cannot grow in lactose. The enzyme transacetylase (30,000 MW) is a product of
lac A whose no definite role has been assigned.
The
lac operon consists of a promoter (P) and an operator (O) together with the structural genes. The initiation codon of
lacZ is TAC that corresponds to AUG of mRNA. It is situated 10 bp away from the end of operator gene. However, the
lac operon cannot functioning in the presence of sugars other than lactose.
(ii) The operator gene. The operator gene is about 28 bp in length present adjacent to
lacZ gene. The base pairs in the operator region are palindrome
i.e. show two-fold symmetry from a point (Fig. 2.11) (Gilbert and Maxam,1973). The operator overlaps the promoter region. The
lac repressor proteins (a tetramer of four subunits) bind to the
lac operator
in vitro and protect part of the promoter region from the digestion of DNase. The repressor proteins bind to the operator and form an operator-repressor complex which in turn physically blocks the transcription of Z,Y and A genes by preventing the release of RNA polymerase to begin transcription (Maniatis and Ptashne,1976).
(iii) The promoter gene. The promoter gene is about 100 nucleotide long and continuous with the operator gene. Gilbert
et al (1974) and Dickson
et al. (1975) have worked out the complete nucleotide sequence of the control region of
lac operon. The promoter gene lies between the operator gene and regulator gene. Like operators the promoter region consists of palindromic sequence of nucleotides. These palindromic sequences are recognized by such proteins that have symmetrically arranged subunits.
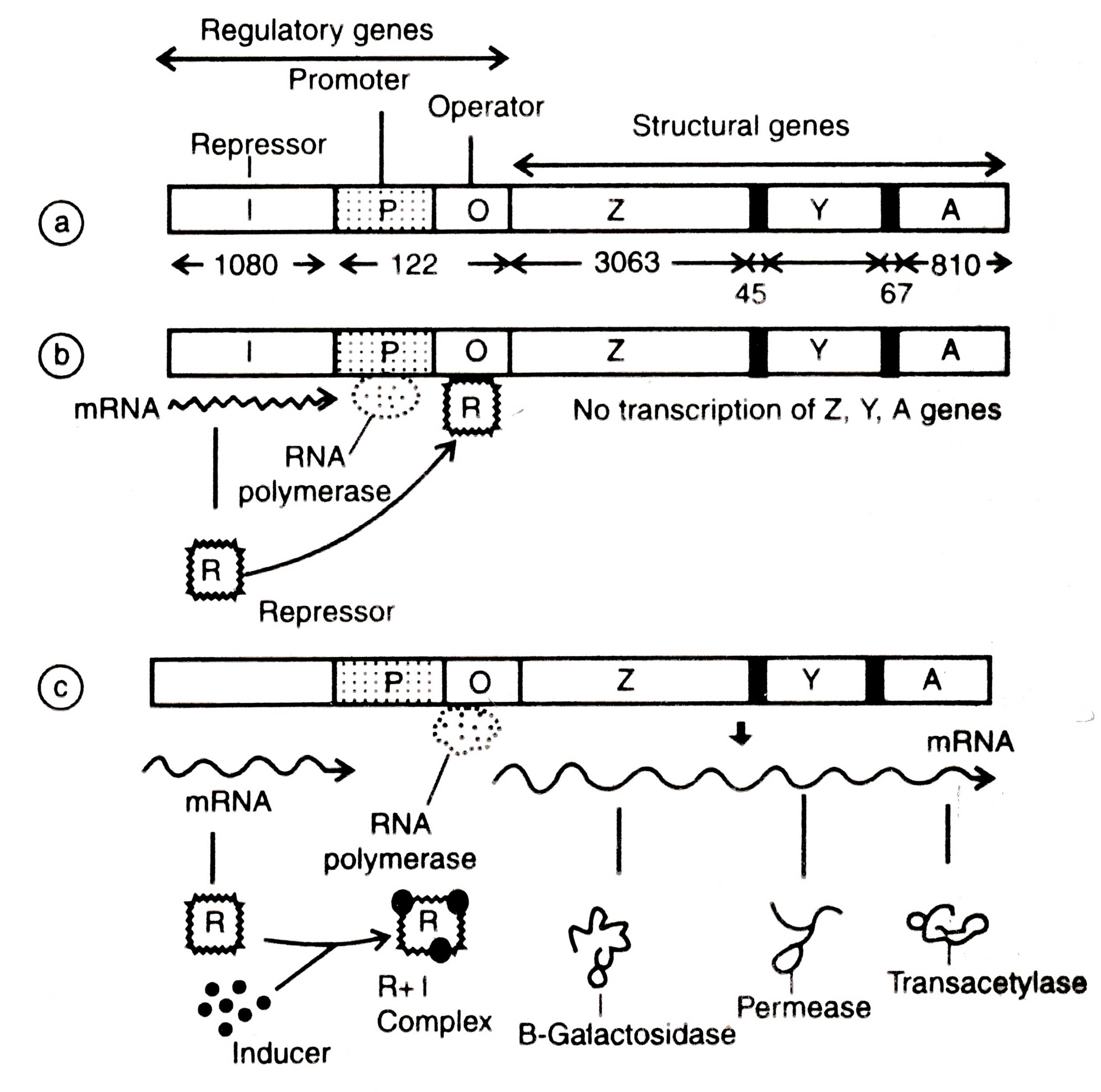
Fig. 2.11. The lac operon; (a) genetic map (the numbers show the number of base pairs that comprise each gene); (b) repressed stat; (c) induced state.
This section of two fold symmetry is present on the CRP site that binds to a protein called CRP (cyclic AMP receptor protein). The CRF is encoded by CRP gene. It has been shown experimentally that CRP binds to cAMP (cyclic AMP found in
E. coli and other organisms) molecule and form a cAMP-CRP complex. This complex is required for transcription because it binds to promoter and enhances the attachment of RN A polymerase to the promoter. Therefore, it increases transcription and translation processes. Thus, cAMP-CRP is a positive regulator in contrast with the repressor, and the
lac operon is controlled both positively and negatively.
According to a model proposed by Pribnow (1975) the promoter region consists of three important components which are present at a fixed position to each other. These components are
(i) the recognition sequence,
(ii) the binding sequence, and
(iii) an mRNA initiation site. The recognition sequence is situated outside the polymerase binding site that is why it is protected from DNase. Firstly, RNA polymerase binds to DNA and forms a complex with the recognition sequence. The binding site is 7 bp long (5'TATGTTG) and present at such region that is protected from DNase. In other organisms the base pairs do not differ from more than two bases. Hence, it can be written as 5' TATPuATG. The mRNA initiation site is present near the binding site on one of the two bases. The initiation site is also protected from DNase. However, there is overlapping of promoter and operator in
lac operon. Moreover, there is a sequence 5'CCGG, 20 bp left to mRNA initiation site. This is known as
Hpall site (5'CCGG) because of being cleaved at this site by the restriction enzyme
HpaII
.
(iv) The repressor (regulator) gene. Repressor gene determines the transcription of structural gene. It is of two types: active and inactive repressors. It codes for amino acid of a defined repressor protein. After synthesis the repressor molecules are diffused from the ribosome and bind to the operator in the absence of an inducer. Finally the path of RNA polymerase is blocked and mRNA is not transcribed. Consequently no protein synthesis occurs. This type of mechanism occurs in the inducible system of active repressor.
Moreover, when an inducer
(e.g. lactose) is present it binds to repressor proteins and forms an inducer-repressor complex. This complex cannot bind to the operator. Due to formation of complex the repressor undergoes changes in conformation of* shape and becomes inactive. Consequently the structural genes can synthesize the polycistronic mRNAs and the later synthesizes enzymes (proteins). In contrast, in the reversible system the regulator gene synthesizes repressor protein that is inactive and, therefore fails to bind to operator. Consequently, proteins are synthesized by the structural genes. However, the repressor proteins can be activated in the presence of a corespressor. The corepressor together with repressor proteins forms the repressor-corepressor complex. This complex binds to operator gene and blocks protein synthesis.
Jacob and Monod (1961) could not identify the repressor protein. Gilbert and Muller - Hill (1966) succeeded in isolating the
lac repressor from the
lac mutant cells of
E. coli inside which the
lac repressor was about ten times greater than the normal cells. The
lac repressor proteins have been crystallized. It has a molecular weight of about 1,50,000 daltons. It consists of four subunits each has 347 amino acid residues and molecular weight of about 40,000 daltons. The repressor proteins have strong affinity for a segment of 12-15 base pairs of operator gene. This binding of repressor blocks the synthesis of mRNA transcript by RNA polymerase.




