Eukaryote / Eukarya
Cell division in eukaryotes is different from organisms without a nucleus (prokaryotes). It involves separating the duplicated chromosomes, through movements directed by microtubules. There are two types of division processes. In mitosis, one cell divides to produce two genetically identical cells. In meiosis, which is required in sexual reproduction, one diploid cell (having two instances of each chromosome, one from each parent) undergoes recombination of each pair of parental chromosomes, and then two stages of cell division, resulting in four haploid cells (gametes). Each gamete has just one complement of chromosomes, each a unique mix of the corresponding pair of parental chromosomes.
Eukaryotes appear to be monophyletic, and so make up one of the three domains of life. The two other domains, bacteria and archaea, are prokaryotes, and have none of the above features.
Contents
» Cell features» Internal membrane
» Mitochondria and plastids
» Cytoskeletal structures
» Plant cell wall
» Differences between eukaryotic cells
» Animal cell
» Plant cell
» Fungal cell
» Other eukaryotic cells
» Reproduction
» Origin and evolution
» See also
» References
 Ostreococcus is the smallest known free living eukaryote with an average size of 0.8 µm.
Ostreococcus is the smallest known free living eukaryote with an average size of 0.8 µm.Domain: Eukaryota
Whittaker & Margulis,1978
Kingdoms
Animalia - Animals
Fungi
Amoebozoa
Plantae - Plants
Chromalveolata
Rhizaria
Excavata
Alternative phylogeny
» Unikonta » Opisthokonta
» Metazoa
» Mesomycetozoa
» Choanozoa
» Eumycota
» Amoebozoa
» Bikonta
» Apusozoa
» Rhizaria
» Excavata
» Archaeplastida
» Chromalveolata
Cell features
Eukaryotic cells are typically much larger than prokaryotes. They have a variety of internal membranes and structures, called organelles, and a cytoskeleton composed of microtubules, microfilaments, and intermediate filaments, which play an important role in defining the cell's organization and shape. Eukaryotic DNA is divided into several linear bundles called chromosomes, which are separated by a microtubular spindle during nuclear division.Internal membrane
Eukaryotic cells include a variety of membrane-bound structures, collectively referred to as the endomembrane system. Simple compartments, called vesicles or vacuoles, can form by budding off other membranes. Many cells ingest food and other materials through a process of endocytosis, where the outer membrane invaginates and then pinches off to form a vesicle. It is probable that most other membrane-bound organelles are ultimately derived from such vesicles.The nucleus is surrounded by a double membrane (commonly referred to as a nuclear envelope), with pores that allow material to move in and out. Various tube- and sheet-like extensions of the nuclear membrane form what is called the endoplasmic reticulum or ER, which is involved in protein transport and maturation. It includes the rough ER where ribosomes are attached, and the proteins they synthesize enter the interior space or lumen. Subsequently, they generally enter vesicles, which bud off from the smooth ER.
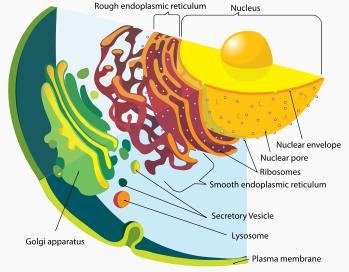 Detail of the endomembrane system and its components
Detail of the endomembrane system and its componentsVesicles may be specialized for various purposes. For instance, lysosomes contain enzymes that break down the contents of food vacuoles, and peroxisomes are used to break down peroxide, which is toxic otherwise. Many protozoa have contractile vacuoles, which collect and expel excess water, and extrusomes, which expel material used to deflect predators or capture prey. In multicellular organisms, hormones are often produced in vesicles. In higher plants, most of a cell's volume is taken up by a central vacuole, which primarily maintains its osmotic pressure.
Mitochondria and plastids
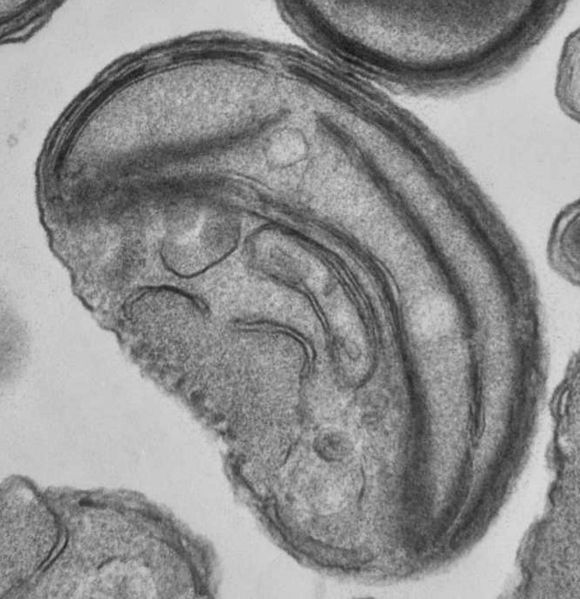
Mitochondria are organelles found in nearly all eukaryotes. They are surrounded by double membranes (known as the phospholipid bi-layer), the inner of which is folded into invaginations called cristae, where aerobic respiration takes place. Mitochondria contain their own DNA and ribosomes and are only formed by the fission of other mitochondria. They are now generally held to have developed from endosymbiotic prokaryotes, probably proteobacteria. The few protozoa that lack mitochondria have been found to contain mitochondrion-derived organelles, such as hydrogenosomes and mitosomes. Plants and various groups of algae also have plastids. Again, these have their own DNA and developed from endosymbiotes, in this case cyanobacteria. They usually take the form of chloroplasts, which like cyanobacteria contain chlorophyll and produce energy through photosynthesis. Others are involved in storing food. Although plastids likely had a single origin, not all plastid-containing groups are closely related. Instead, some eukaryotes have obtained them from others through secondary endosymbiosis or ingestion.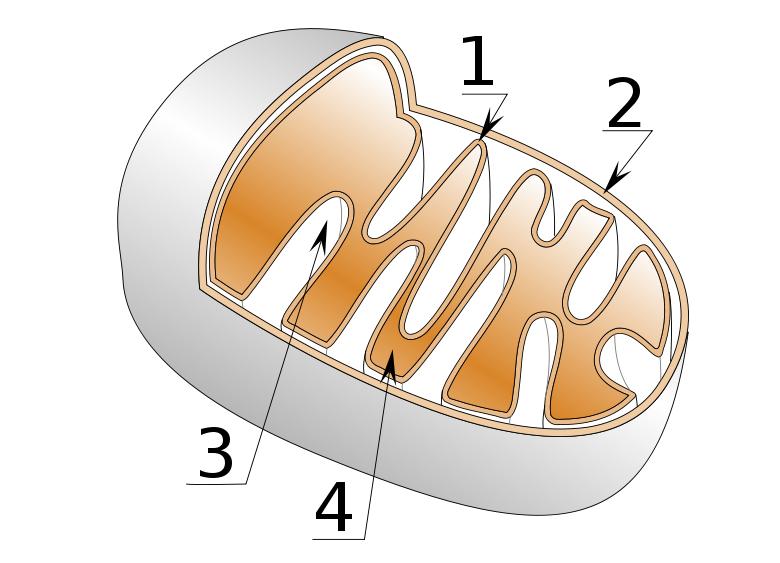 Mitochondria structure:
Mitochondria structure:1) Inner membrane
2) Outer membrane
3) Crista
4) Matrix
Cytoskeletal structures
Many eukaryotes have long slender motile cytoplasmic projections, called flagella, or similar, but shorter structures called cilia. Flagella and cilia are sometimes referred to as undulipodia, and are variously involved in movement, feeding, and sensation. They are composed mainly of tubulin. These are entirely distinct from prokaryotic flagella. They are supported by a bundle of microtubules arising from a basal body, also called a kinetosome or centriole, characteristically arranged as nine doublets surrounding two singlets. Flagella also may have hairs, or mastigonemes, and scales connecting membranes and internal rods. Their interior is continuous with the cell's cytoplasm.Centrioles are often present even in cells and groups that do not have flagella. They generally occur in groups of one or two, called kinetids, that give rise to various microtubular roots. These form a primary component of the cytoskeletal structure, and are often assembled over the course of several cell divisions, with one flagellum retained from the parent and the other derived from it. Centrioles may also be associated in the formation of a spindle during nuclear division.
Significance of cytoskeletal structures is underlined in determination of shape of the cells, as well as their being essential components of migratory responses like chemotaxis and chemokinesis. Some protists have various other microtubule-supported organelles. These include the radiolaria and heliozoa, which produce axopodia used in flotation or to capture prey, and the haptophytes, which have a peculiar flagellum-like organelle called the haptonema.
Plant cell wall
Plant cells have a cell wall, a fairly rigid layer outside the cell membrane, providing the cell with structural support, protection, and a filtering mechanism. The cell wall also prevents over-expansion when water enters the cell. The major carbohydrates making up the primary cell wall of land plants are cellulose, hemicellulose, and pectin. The cellulose microfibrils are linked via hemicellulosic tethers to form the cellulose-hemicellulose network, which is embedded in the pectin matrix. The most common hemicellulose in the primary cell wall is xyloglucan.Differences between eukaryotic cells
There are many different types of eukaryotic cells, though animals and plants are the most familiar eukaryotes, and thus provide an excellent starting point for understanding eukaryotic structure. Fungi and many protists have some substantial differences, however.Animal cell
An animal cell is a form of eukaryotic cell that makes up many tissues in animals. The animal cell is distinct from other eukaryotes, most notably plant cells, as they lack cell walls and chloroplasts, and they have smaller vacuoles. Due to the lack of a rigid cell wall, animal cells can adopt a variety of shapes, and a phagocytic cell can even engulf other structures. There are many different cell types. For instance, there are approximately 210 distinct cell types in the adult human body.Plant cell
Plant cells are quite different from the cells of the other eukaryotic organisms. Their distinctive features are:» A large central vacuole (enclosed by a membrane, the tonoplast), which maintains the cell's turgor and controls movement of molecules between the cytosol and sap
» A primary cell wall containing cellulose, hemicellulose and pectin, deposited by the protoplast on the outside of the cell membrane; this contrasts with the cell walls of fungi, which contain chitin, and the cell envelopes of prokaryotes, in which peptidoglycans are the main structural molecules
» The plasmodesmata, linking pores in the cell wall that allow each plant cell to communicate with other adjacent cells; this is different from the functionally analogous system of gap junctions between animal cells.
» Plastids, especially chloroplasts that contain chlorophyll, the pigment that gives plants their green color and allows them to perform photosynthesis
» Higher plants, including conifers and flowering plants (Angiospermae) lack the flagellae and centrioles that are present in animal cells.
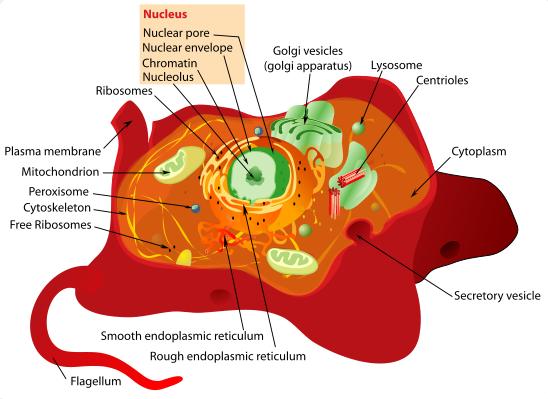
Structure of a typical animal cell.
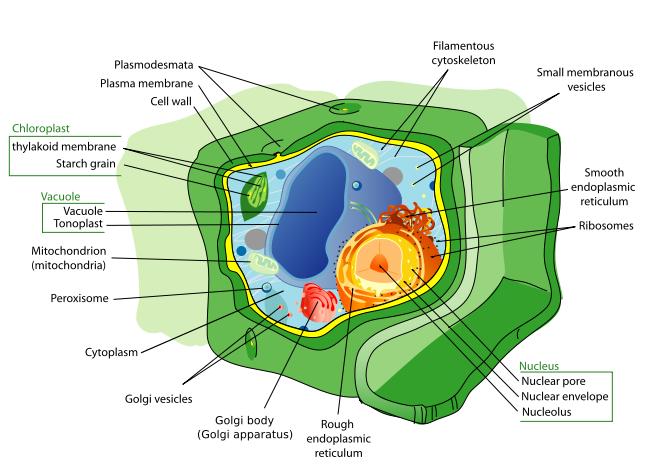
Structure of a typical plant cell.
Fungal cell
Fungal cells are most similar to animal cells, with the following exceptions: » A cell wall containing chitin» Less definition between cells; the hyphae of higher fungi have porous partitions called septa, which allow the passage of cytoplasm, organelles, and, sometimes, nuclei. Primitive fungi have few or no septa, so each organism is essentially a giant multinucleate supercell; these fungi are described as coenocytic.
» Only the most primitive fungi, chytrids, have flagella.
Other eukaryotic cells
Eukaryotes are a very diverse group, and their cell structures are equally diverse. Many have cell walls; many do not. Many have chloroplasts, derived from primary, secondary, or even tertiary endosymbiosis; and many do not. Some groups have unique structures, such as the cyanelles of the glaucophytes, the haptonema of the haptophytes, or the ejectisomes of the cryptomonads. Other structures, such as pseudopods, are found in various eukaryote groups in different forms, such as the lobose amoebozoans or the reticulose foraminiferans.Reproduction
Nuclear division is often coordinated with cell division. This generally takes place by mitosis, a process that allows each daughter nucleus to receive one copy of each chromosome. In most eukaryotes, there is also a process of sexual reproduction, typically involving an alternation between haploid generations, wherein only one copy of each chromosome is present, and diploid generations, wherein two are present, occurring through nuclear fusion (syngamy) and meiosis. There is considerable variation in this pattern, however. Eukaryotes have a smaller surface to volume area ratio than prokaryotes, and thus have lower metabolic rates and longer generation times. In some multicellular organisms, cells specialized for metabolism will have enlarged surface areas, such as intestinal vili.Origin and evolution
The origin of the eukaryotic cell was a milestone in the evolution of life, since they include all complex cells and almost all multi-cellular organisms. The timing of this series of events is hard to determine; Knoll (2006) suggests they developed approximately 1.6 - 2.1 billion years ago. Some acritarchs are known from at least 1650 million years ago, and the possible alga Grypania has been found as far back as 2100 million years ago. Fossils that are clearly related to modern groups start appearing around 1.2 billion years ago, in the form of a red alga.Biomarkers suggest that at least stem eukaryotes arose even earlier. The presence of steranes in Australian shales indicates that eukaryotes were present 2.7 billion years ago.
rRNA trees constructed during the 1980s and 1990s left most eukaryotes in an unresolved "crown" group (not technically a true crown), which was usually divided by the form of the mitochondrial cristae; see crown eukaryotes. The few groups that lack mitochondria branched separately, and so the absence was believed to be primitive; but this is now considered an artifact of long-branch attraction, and they are known to have lost them secondarily.
More recent work has painted a different picture. Most eukaryotes are now included in one of the following supergroups, although the relationship between these groups, and the monophyly of each group, is not yet clear:
Opisthokonts
Animals, fungi, choanoflagellates, etc.
Amoebozoa
Most lobose amoebae and slime moulds
Rhizaria
Foraminifera, Radiolaria, and various other amoeboid protozoa
Excavates
Various flagellate protozoa
Archaeplastida (or Primoplantae)
Land plants, green algae, red algae, and glaucophytes
Chromalveolates
Heterokonts, Haptophytes, Cryptomonads, and Alveolates.
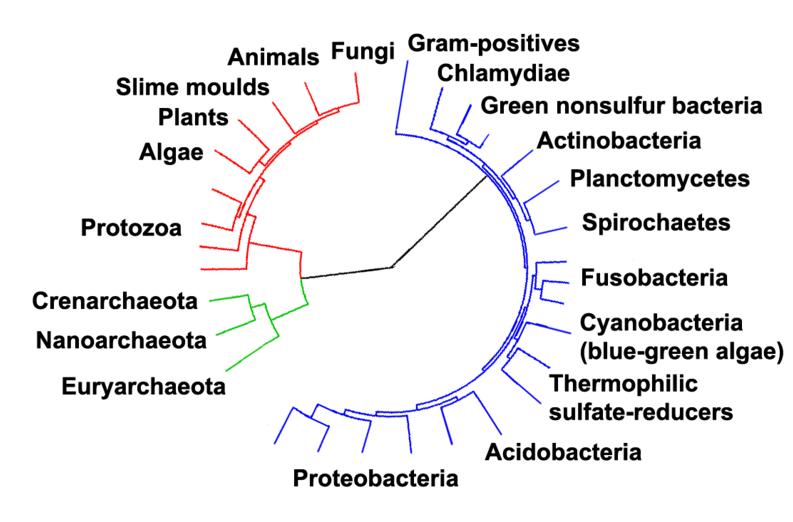
Phylogenetic tree showing the relationship between the eukaryotes and other forms of life. Eukaryotes are colored red, archaea green and bacteria blue.
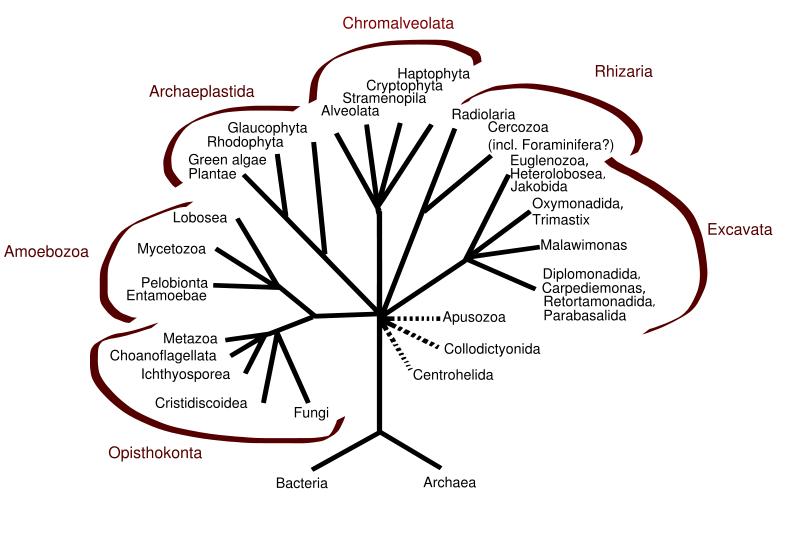
One hypothesis of eukaryotic relationships
A further proposal is that eukaryotes can be classified into two larger clades, the unikonts and the bikonts, that derive from an ancestral uniflagellar organism and a biflagellate, respectively. In this system, the opisthokonts and amoebozoans are considered unikonts, and the rest bikonts. The correct relationships of the organisms in the Archaeplastida and Chromalveolates is far from clear, and it seems likely that neither group, as defined above, is monophyletic. Some small protist groups have not been related to any of these supergroups, in particular the characteristics of the anaerobic protist Breviata anathema seems to contradict the unikont/bikont division.
- Eukaryotes resulted from the complete fusion of two or more cells, wherein the cytoplasm formed from a eubacterium, and the nucleus from an archaeon, or from a virus.
- Eukaryotes developed from Archaea, and acquired their eubacterial characteristics from the proto-mitochondrion.
- Eukaryotes and Archaea developed separately from a modified eubacterium.
In a study using genomes to construct supertrees, Pisani et al. (2007) suggest that, along with evidence that there was never a mitochondrion-less eukaryote, eukaryotes evolved from a syntrophy between an archaea closely related to Thermoplasmatales and an α-proteobacterium, likely a symbiosis driven by sulfur or hydrogen. The mitochondrion and its genome is a remnant of the α-proteobacterial endosymbiont.
See also
» ProkaryoteReferences
- Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P (2006). "Toward automatic reconstruction of a highly resolved tree of life". Science 311 (5765): 1283–7.- Knoll, Andrew H.; Javaux, E.J, Hewitt, D. and Cohen, P. (2006). "Eukaryotic organisms in Proterozoic oceans". Philosophical Transactions of the Royal Society of London, Part B 361 (1470): 1023–1038.
- Archean Molecular Fossils and the Early Rise of Eukaryotes, by Jochen Brocks et al., Science, 13 Aug. 1999, pp. 1033-6.
- Mass extinctions: the microbes strike back by Peter Ward, New Scientist, 9 Feb. 2008, pp. 40-3.
- Tovar J, Fischer A, Clark CG (1999). "The mitosome, a novel organelle related to mitochondria in the amitochondrial parasite Entamoeba histolytica". Mol. Microbiol. 32 (5): 1013–21.
- Boxma B, de Graaf RM, van der Staay GW, et al (2005). "An anaerobic mitochondrion that produces hydrogen". Nature 434 (7029): 74–9.
- Burki F, Shalchian-Tabrizi K, Minge M, Skjæveland Å, Nikolaev SI, et al. (2007). "Phylogenomics Reshuffles the Eukaryotic Supergroups". PLoS ONE 2 (8: e790): e790.
- Burki F, Pawlowski J (October 2006). "Monophyly of Rhizaria and multigene phylogeny of unicellular bikonts". Mol. Biol. Evol. 23 (10): 1922–30.
- Kim, E; Graham, Le (Jul 2008). "EEF2 analysis challenges the monophyly of Archaeplastida and Chromalveolata." (Free full text). PLoS ONE 3 (7): e2621.
- Roger AJ, Simpson AGB. (2009). "Evolution: Revisiting the Root of the Eukaryote Tree". Current Biology 19 (4): R165-R167.
- Martin W (December 2005). "Archaebacteria (Archaea) and the origin of the eukaryotic nucleus". Curr. Opin. Microbiol. 8 (6): 630–7.
- Takemura M (May 2001). "Poxviruses and the origin of the eukaryotic nucleus". J. Mol. Evol. 52 (5): 419–25.
- Bell PJ (September 2001). "Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?". J. Mol. Evol. 53 (3): 251–6.
- Jékely G (2007). "Origin of eukaryotic endomembranes: a critical evaluation of different model scenarios". Adv. Exp. Med. Biol. 607: 38–51.
- Cavalier-Smith T (March 2002). "The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa". Int. J. Syst. Evol. Microbiol. 52 (Pt 2): 297–354.
- Martin W, Müller M (March 1998). "The hydrogen hypothesis for the first eukaryote". Nature 392 (6671): 37–41.
- Pisani D, Cotton JA, McInerney JO (2007). "Supertrees disentangle the chimerical origin of eukaryotic genomes". Mol Biol Evol. 24 (8): 1752–60.
Support our developers




