Prokaryote
The prokaryotes are divided into two domains: the bacteria and the archaea. Archaea were recognized as a domain of life in 1990. These organisms were originally thought to live only in inhospitable conditions such as extremes of temperature, pH, and radiation but have since been found in all types of habitats.
Contents
» Relationship to eukaryotes» Sociality
» Reproduction
» Structure
» Morphology of prokaryotic cells
» Environment
» Evolution of prokaryotes
» References
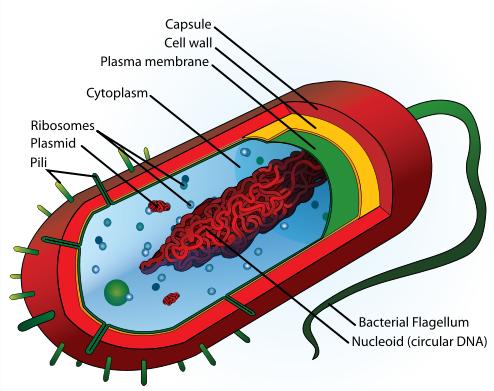
Cell structure of a bacterium, one of the two groups of prokaryotic life.
Relationship to eukaryotes
A distinction between prokaryotes and eukaryotes (meaning true kernel, also spelled "eucaryotes") is that eukaryotes do have "true" nuclei containing their DNA, whereas the genetic material in prokaryotes is not membrane-bound. Eukaryotic organisms may be unicellular, as in amoebae, or multicellular, as in plants and humans. The difference between the structure of prokaryotes and eukaryotes is so great that it is sometimes considered to be the most important distinction among groups of organisms. However, a criticism of this classification is that the word "prokaryote" is based on what these organisms are not (they are not eukaryotic), rather than what they are (either archaea or bacteria). In 1977, Carl Woese proposed dividing prokaryotes into the Bacteria and Archaea (originally Eubacteria and Archaebacteria) because of the major differences in the structure and genetics between the two groups of organisms. This arrangement of Eukaryota (also called "Eukarya"), Bacteria, and Archaea is called the three-domain system replacing the traditional two-empire system.Prokaryotes have a larger surface area to volume ratio giving them a higher metabolic rate, a higher growth rate and consequently a shorter generation time compared to Eukaryotes.
Sociality
While prokaryotes are still commonly imagined to be strictly unicellular, most are capable of forming stable aggregate communities. When such communities are encased in a stabilizing polymer matrix (“slime”), they may be called “biofilms”. Cutting edge research shows that, like those in multicellular organisms, cells in biofilms often show distinct patterns of gene expression (phenotypic differentiation) in time and space. Also, like multicellular eukaryotes, these changes in expression appear to often result from cell-to-cell signaling, a phenomenon known as quorum sensing.Biofilms may be highly heterogeneous and structurally complex and may attach to solid surfaces, or exist at liquid-air interfaces, or potentially even liquid-liquid interfaces. Bacterial biofilms are often comprised of microcolonies (approximately dome-shaped masses of bacteria and matrix) separated by “voids” through which the medium (e.g. water) may flow relatively uninhibited. The microcolonies may join together above the substratum to form a continuous layer, closing the network of channels separating microcolonies. This structural complexity—combined with observations that oxygen limitation (a ubiquitous challenge for anything growing in size beyond the scale of diffusion) is at least partially eased by movement of medium throughout the biofilm—has led some to speculate that this may constitute a circulatory system.
Reproduction
Bacteria and archaea reproduce through asexual reproduction, usually by binary fission or budding. Genetic exchange and recombination still occur, but this is a form of horizontal gene transfer and is not a replicative process, simply involving DNA being transferred between two cells, as in bacterial conjugation.Structure
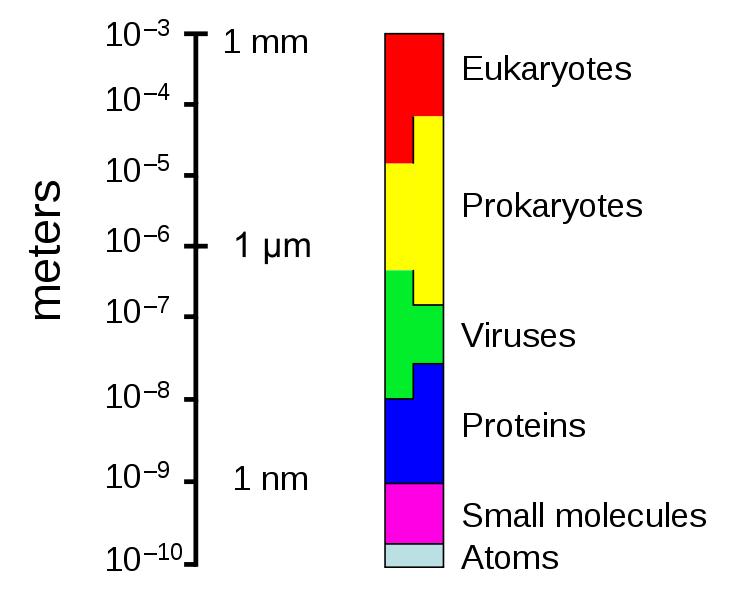
The sizes of prokaryotes relative to other organisms and biomolecules
Procaryotes vary in size between 0.2µm and 700µm (Thiomargarita namibiensis up to 750µm).
| Prokaryotic cell Structure |
|---|
| Flagellum |
| Cell membrane |
| Cell wall (except genus Mycoplasma) |
| Cytoplasm |
| Ribosome |
| Nucleoid |
| Glycocalyx |
| Inclusions |
Morphology of prokaryotic cells
Prokaryotic cells have various shapes; the four basic shapes are:» Cocci - spherical
» Bacilli - rod shaped
» Spirochaete - spiral shaped
» Vibrio - comma shaped
Environment
Prokaryotes live in nearly all environments on earth where there is liquid water. Some archaea and bacteria thrive in harsh conditions, such as high temperatures (thermophiles) or high salinity (halophiles). Organisms such as these are referred to as extremophiles. Many archaea grow as plankton in the oceans. Symbiotic prokaryotes live in or on the bodies of other organisms, including humans.Evolution of prokaryotes
It is generally accepted that the first living organisms were some form of prokaryotes, which may have evolved out of protobionts. The oldest known fossilized prokaryotes were laid down approximately 3.5 billion years ago, only about 1 billion years after the formation of the earth's crust. Even today, prokaryotes are perhaps the most successful and abundant life forms. Eukaryotes only formed later, from endosymbiosis of multiple prokaryote ancestors. The oldest known fossil eukaryotes are about 1.7 billion years old. However, some genetic evidence suggests eukaryotes appeared as early as 3 billion years ago.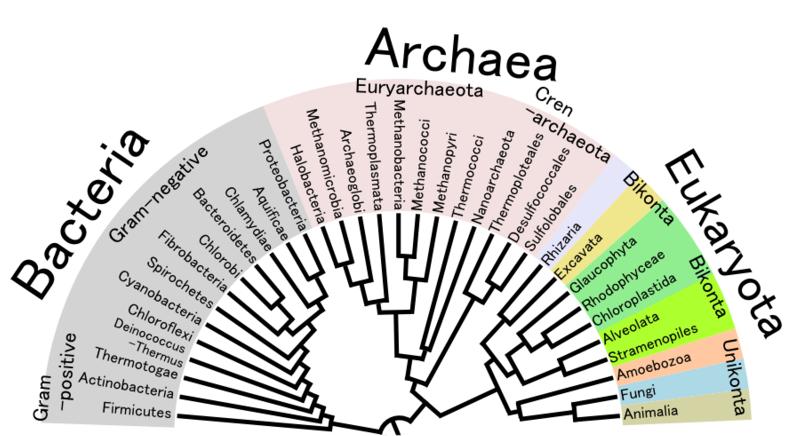
Phylogenetic tree showing the diversity of prokaryotes, compared to eukaryotes.
Prokaryotes have diversified greatly throughout their long existence. The metabolism of prokaryotes is far more varied than that of eukaryotes, leading to many highly distinct prokaryotic types. For example, in addition to using photosynthesis or organic compounds for energy, as eukaryotes do, prokaryotes may obtain energy from inorganic compounds such as hydrogen sulfide. This enables prokaryotes to thrive in harsh environments as cold as the snow surface of Antarctica, and as hot as undersea hydrothermal vents and land-based hot springs.
References
- Kaiser D (October 2003). "Coupling cell movement to multicellular development in myxobacteria". Nat. Rev. Microbiol. 1 (1): 45–54.- Sapp J (June 2005). "The prokaryote-eukaryote dichotomy: meanings and mythology". Microbiol. Mol. Biol. Rev. 69 (2): 292–305.
- Woese CR (March 1994). "There must be a prokaryote somewhere: microbiology's search for itself". Microbiol. Rev. 58 (1): 1–9.
- Thanbichler M, Wang S, Shapiro L (2005). "The bacterial nucleoid: a highly organized and dynamic structure". J Cell Biochem 96 (3): 506–21.
- Harold F (1972). "Conservation and transformation of energy by bacterial membranes". Bacteriol Rev 36 (2): 172–230.
- Shih YL, Rothfield L (2006). "The bacterial cytoskeleton". Microbiol. Mol. Biol. Rev. 70 (3): 729–54.
- Michie KA, Löwe J (2006). "Dynamic filaments of the bacterial cytoskeleton". Annu. Rev. Biochem. 75: 467–92.
- Costeron, J. W., Lewandowski, Z., Caldwell, D. E., Korber, D. R., and Lappin-Scott, H. M. 1995. “Microbial biofilms.” Annu. Rev. Microbiol. 49: 711-45.
- Shapiro, J. A. 1998. “Thinking about bacterial populations as multicellular organisms.” Annu. Rev. Microbiol. 52: 81-104.
- Hamilton, W. D. 1964. “The genetical evolution of social behavior. II.” J. Theor. Biol.7:17-52.
- Costerton, J. W., Stewart, P. S., and Greenberg, E. P. 1999. “Bacterial biofilms: a common cause of persistent infections.” Science 284: 1318-1322.
- Carl Woese, J Peter Gogarten, "When did eukaryotic cells (cells with nuclei and other internal organelles) first evolve? What do we know about how they evolved from earlier life-forms?" Scientific American, October 21, 1999.
- McSween HY (1997). "Evidence for life in a martian meteorite?". GSA Today 7 (7): 1–7.
- McKay D. S., Gibson E. K., ThomasKeprta K. L., Vali H., Romanek C. S., Clemett S. J., Chillier X. D. F., Maechling C. R., Zare R. N. (1996). "Search for past life on Mars: Possible relic biogenic activity in Martian meteorite ALH84001". Science 273: 924–930.
- Scott ER (1999). "Origin of carbonate-magnetite-sulfide assemblages in Martian meteorite ALH84001". J. Geophys. Res. 104 (E2): 3803–13.
Support our developers




