Class Asteroidea
Class Asteroidea
Sea stars, often called starfishes, demonstrate the basic features of echinoderm structure and function very well, and they are easily obtainable. Thus we will consider them first, then comment on major differences shown by other groups.
Sea stars are familiar along the shoreline where large numbers may aggregate on rocks. Sometimes they cling so tenaciously that they are difficult to dislodge without tearing off some tube feet. They also live on muddy or sandy bottoms and among coral reefs. They are often brightly colored and range in size from a centimeter in greatest diameter to about a meter across. Asterias (Gr. asteros, a star) is one of the common genera of the east coast of the United States and is commonly studied in zoology laboratories. Pisaster (Gr. pisos, a pea, + asteros, a star) is common on the west coast of the United States, as is Dermasterias (Gr. dermatos, skin, leather, + asteros, a star), the leather star.
Form and Function
External Features
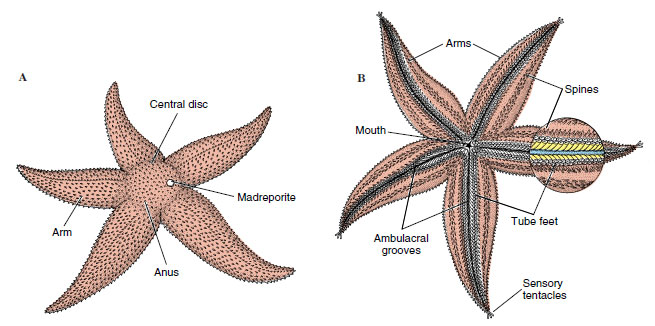
Sea stars are composed of a central disc that merges gradually with the tapering arms (rays). The body is somewhat flattened, flexible, and covered with a ciliated, pigmented epidermis. The mouth is centered on the under, or oral, side, surrounded by a soft peristomial membrane. An ambulacrum (pl., ambulacra, L. ambulacrum, a covered way, an alley, a walk planted with trees) or ambulacral area, runs from the mouth on the oral side of each arm to the tip of the arm. Sea stars typically have five arms, but they may have more (Figure 23-1D), and there are as many ambulacral areas as there are arms. An ambulacral groove is found along the middle of each ambulacral area, and the groove is bordered by rows of tube feet (podia) (Figure 23-2). These in turn are usually protected by movable spines. A large radial nerve can be seen in the center of each ambulacral groove (Figure 23-3C), between the rows of tube feet. The nerve is very superficially located, covered only by thin epidermis. Under the nerve is an extension of the coelom and the radial canal of the water-vascular system, all of which are external to the underlying ossicles (Figure 23-3C). In all other classes of living echinoderms except crinoids, these structures are covered by ossicles or other dermal tissue; thus ambulacral grooves in asteroids and crinoids are said to be open, and those of the other groups are closed.
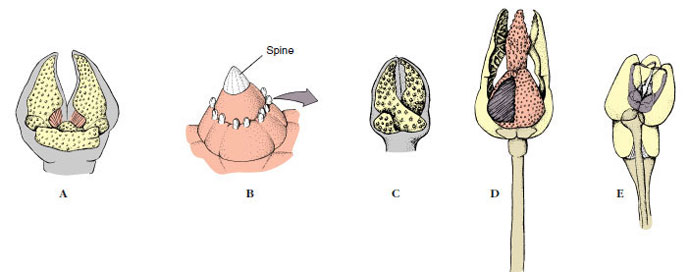
The aboral surface is usually rough and spiny, although spines of many species are flattened, so that the surface appears smooth (Figure 23-1C). Around the bases of spines are groups of minute, pincerlike pedicellariae, bearing tiny jaws manipulated by muscles (Figure 23-4). These jaws help keep the body surface free of debris, protect the papulae, and sometimes aid in food capture. Papulae (dermal branchiae or skin gills) are soft delicate projections of the coelomic cavity, covered only with epidermis and lined internally with peritoneum; they extend out through spaces between ossicles and are concerned with respiration (Figures 23-3C, and 23-4F). Also on the aboral side are the inconspicuous anus and the circular madreporite (Figure 23-2A), a calcareous sieve leading to the water-vascular system.
Endoskeleton
Beneath the epidermis of sea stars is a mesodermal endoskeleton of small calcareous plates, or ossicles, bound together with connective tissue. From these ossicles project the spines and tubercles that make up the spiny surface. Ossicles are penetrated by a meshwork of spaces, usually filled with fibers and dermal cells. This internal meshwork structure is described as stereom and is unique to echinoderms.
Muscles in the body wall move the rays and can partially close the ambulacral grooves by drawing their margins together.
Coelom, Excretion, and Respiration
The coelomic compartments of larval echinoderms give rise to several structures in adults, one of which is a spacious body coelom filled with fluid. The fluid contains amebocytes (coelomocytes), bathes the internal organs, and projects into the papulae. The ciliated peritoneal lining of the coelom circulates the fluid around the body cavity and into the papulae. Exchange of respiratory gases and excretion of nitrogenous waste, principally ammonia, take place by diffusion through the thin walls of papulae and tube feet. Some wastes may be picked up by coelomocytes, which migrate through the epithelium of the papulae or tube feet to the exterior, or the tips of papulae containing waste-laden coelomocytes may pinch off.
Water-Vascular System
The water-vascular system is another coelomic compartment and is unique to echinoderms. Showing exploitation of hydraulic mechanisms to a greater degree than in any other animal group, it is a system of canals and specialized tube feet that, together with the dermal ossicles, has determined the evolutionary potential and limitations of this phylum. In sea stars the primary functions of the water-vascular system are locomotion and food gathering, in addition to respiration and excretion.
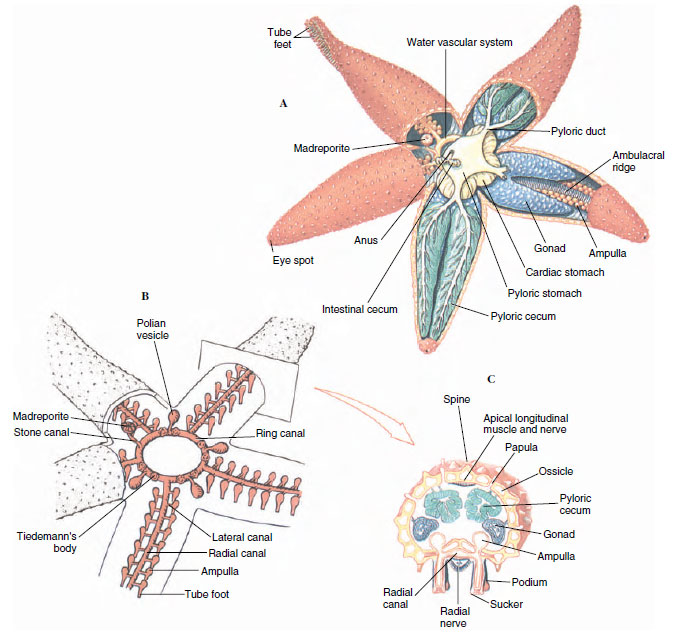
Structurally, the water-vascular system opens to the outside through small pores in the madreporite. The madreporite of asteroids is on the aboral surface (Figure 23-2A) and leads into the stone canal, which descends toward the ring canal around the mouth (Figure 23-3B). Radial canals diverge from the ring canal, one into the ambulacral groove of each ray. Also attached to the ring canal are four or five pairs of folded, pouchlike Tiedemann’s bodies and from one to five polian vesicles (polian vesicles are absent in some sea stars, such as Asterias). Tiedemann’s bodies may produce coelomocytes, and polian vesicles are apparently for fluid storage.
A series of small lateral canals, each with a one-way valve, connects the radial canal to the cylindrical podia, or tube feet, along the sides of the ambulacral groove in each ray. Each podium is a hollow, muscular tube, the inner end of which is a muscular sac, or ampulla, that lies within the body coelom (Figure 23-3A and C), and the outer end of which usually bears a sucker. Some species lack suckers. Podia pass to the outside between ossicles in the ambulacral groove.
The water-vascular system operates hydraulically and is an effective locomotor mechanism. The valves in the lateral canals prevent backflow of fluid into the radial canals. Each tube foot has in its walls connective tissue that maintains the cylinder at a relatively constant diameter. Contraction of muscles in the ampulla forces fluid into the podium, extending it. Conversely, contraction of the longitudinal muscles in the tube foot retracts the podium, forcing fluid back into the ampulla. Contraction of muscles in one side of the podium bends the organ toward that side. Small muscles at the end of the tube foot can raise the middle of the disclike end, creating suction when the end is applied to a firm substrate. It has been estimated that by combining mucous adhesion with suction, a single podium can exert a pull equal to 25 to 30 g. Coordinated action of all or many of the tube feet is sufficient to draw the animal up a vertical surface or over rocks. The ability to move while firmly adhering to the substrate is a clear advantage to an animal living in a sometimes wave-churned environment.
On a soft surface, such as muck or sand, suckers are ineffective (numerous sand-dwelling species have no suckers), so the tube feet are employed as legs. Locomotion becomes mainly a stepping process. Most sea stars can move only a few centimeters per minute, but some very active ones can move 75 to 100 cm per minute; for example, Pycnopodia (Gr. pyknos, compact, dense, + pous, podos, foot) (Figure 23-5B). When inverted, a sea star bends its rays until some of the tubes reach the substratum and attach as an anchor; then it slowly rolls over.
Tube feet are innervated by the central nervous system (ectoneural and hyponeural systems, see following text). Nervous coordination enables tube feet to move in a single direction, although not in unison, so that the sea star may progress. If the radial nerve in an arm is cut, podia in that arm lose coordination, although they can still function. If the circumoral nerve ring is cut, podia in all arms become uncoordinated, and movement ceases.
Feeding and Digestive System
The mouth on the oral side leads through a short esophagus to a large stomach in the central disc. The lower (cardiac) part of the stomach can be everted through the mouth during feeding (Figure 23-2B), and excessive eversion is prevented by gastric ligaments. The upper (pyloric) part is smaller and connects by ducts to a pair of large pyloric ceca (digestive glands) in each arm (Figure 23-3A). Digestion is mostly extracellular, although some intracellular digestion may occur in the ceca. A short intestine leads aborally from the pyloric stomach, and there are usually a few small, saclike intestinal ceca (Figure 23-3A). The anus is inconspicuous, and some sea stars lack an intestine and anus.
Many sea stars are carnivorous and feed on molluscs, crustaceans, polychaetes, echinoderms, other invertebrates, and sometimes small fish. Sea stars consume a wide range of food items, but many show particular preferences (Figures 23-5 and 23-6). Some select brittle stars, sea urchins, or sand dollars, swallowing them whole and later regurgitating undigestible ossicles and spines (Figure 23-5B). Some attack other sea stars, and if they are small compared with their prey, they may attack and begin eating at the end of one arm.
Some asteroids feed heavily on molluscs (Figure 23-5A), and Asterias is a significant predator on commercially important clams and oysters. When feeding on a bivalve, a sea star will hump over its prey, attaching its podia to the valves, and then exert a steady pull, using its feet in relays. A force of some 1300 g can thus be exerted. In half an hour or so the adductor muscles of the bivalve fatigue and relax. With a very small gap available, the star inserts its soft everted stomach into the space between the valves and wraps it around the soft parts of the shellfish. After feeding, the sea star draws its stomach inward by contraction of the stomach muscles and relaxation of body-wall muscles.
Some sea stars feed on small particles, either entirely or in addition to carnivorous feeding. Plankton and other organic particles coming in contact with the animal’s surface are carried by the epidermal cilia to the ambulacral grooves and then to the mouth.
Hemal System
The so-called hemal system is not very well developed in asteroids, and its function in all echinoderms is unclear. The hemal system has little or nothing to do with circulation of body fluids. It is a system of tissue strands enclosing unlined sinuses and is itself enclosed in another coelomic compartment, the perihemal channels (Figure 23-7). The hemal system may be useful in distributing digested products, but its specific functions are not really known.
Nervous System
The nervous system consists of three units at different levels in the disc and arms. Chief of these systems is the oral (ectoneural) system composed of a nerve ring around the mouth and a main radial nerve into each arm. It appears to coordinate the tube feet. A deep (hyponeural) system lies aboral to the oral system, and an aboral system consists of a ring around the anus and radial nerves along the roof of each ray. An epidermal nerve plexus or nerve net freely connects these systems with the body wall and related structures. The epidermal plexus coordinates responses of the dermal branchiae to tactile stimulation—the only instance known in echinoderms in which coordination occurs through a nerve net.
Sense organs are not well developed. Tactile organs and other sensory cells are scattered over the surface, and an ocellus is at the tip of each arm. Their reactions are mainly to touch, temperature, chemicals, and differences in light intensity. Sea stars are usually more active at night.
Reproductive System, Regeneration, and Autotomy
Most sea stars have separate sexes. A pair of gonads lies in each interradial space (Figure 23-3A). Fertilization is external and occurs in early summer when eggs and sperm are shed into the water. A secretion from neurosecretory cells located on the radial nerves stimulates maturation and shedding of asteroid eggs.
Echinoderms can regenerate lost parts. Sea star arms can regenerate readily, even if all are lost. Sea stars also have the power of autotomy and can cast off an injured arm near the base. Regeneration of a new arm may take several months.
Some species can regenerate a complete new sea star (Figure 23-8) from a detached arm that contains a part (about one-fifth) of the central disc. In former times fishermen used to dispatch sea stars they collected from their oyster beds by chopping them in half with a hatchet—a worse than futile activity. Some sea stars reproduce asexually under normal conditions by cleaving the central disc, each part regenerating the rest of the disc and missing arms.
Development
In some species the liberated eggs are brooded, either under the oral side of the animal or in specialized aboral structures, and development is direct, but in most species embryonating eggs are free in the water and hatch to freeswimming larvae.
Early embryogenesis shows a typical primitive deuterostome pattern (see Figures 8-7A and 8-11A). Gastrulation is by invagination, and the anterior end of the archenteron pinches off to become the coelomic cavity, which expands in a U shape to fill the blastocoel. Each of the legs of the U, at the posterior, constricts to become a separate vesicle, and these eventually give rise to the main coelomic compartments of the body (metacoels, called somatocoels in echinoderms). The anterior portion of the U undergoes subdivision to form the protocoels and mesocoels (called axocoels and hydrocoels in echinoderms) (Figure 23-9). The left hydrocoel will become the water-vascular system, and the left axocoel will give rise to the stone canal and perihemal channels. The right axocoel and hydrocoel will disappear. The free-swimming larva has cilia arranged in bands and is called a bipinnaria (Figure 23-10A). These ciliated tracts become extended into larval arms. Soon the larva grows three adhesive arms and a sucker at its anterior end and is then called a brachiolaria. At that time it attaches to the substratum, forms a temporary attachment stalk, and undergoes metamorphosis.
Metamorphosis involves a dramatic reorganization of a bilateral larva into a radial juvenile. The anteroposterior axis of the larva is lost, and what was the left side becomes the oral surface, and the larval right side becomes the aboral surface (Figure 23-9). Correspondingly, the larval mouth and anus disappear, and a new mouth and anus form on what were originally the left and right sides, respectively. The portion of the anterior coelomic compartment from the left side expands to form the ring canal of the watervascular system around the mouth, and then it grows branches to form the radial canals. As the short, stubby arms and the first podia appear, the animal detaches from its stalk and begins life as a young sea star.
Sea stars, often called starfishes, demonstrate the basic features of echinoderm structure and function very well, and they are easily obtainable. Thus we will consider them first, then comment on major differences shown by other groups.
Sea stars are familiar along the shoreline where large numbers may aggregate on rocks. Sometimes they cling so tenaciously that they are difficult to dislodge without tearing off some tube feet. They also live on muddy or sandy bottoms and among coral reefs. They are often brightly colored and range in size from a centimeter in greatest diameter to about a meter across. Asterias (Gr. asteros, a star) is one of the common genera of the east coast of the United States and is commonly studied in zoology laboratories. Pisaster (Gr. pisos, a pea, + asteros, a star) is common on the west coast of the United States, as is Dermasterias (Gr. dermatos, skin, leather, + asteros, a star), the leather star.
Form and Function
External Features
Sea stars are composed of a central disc that merges gradually with the tapering arms (rays). The body is somewhat flattened, flexible, and covered with a ciliated, pigmented epidermis. The mouth is centered on the under, or oral, side, surrounded by a soft peristomial membrane. An ambulacrum (pl., ambulacra, L. ambulacrum, a covered way, an alley, a walk planted with trees) or ambulacral area, runs from the mouth on the oral side of each arm to the tip of the arm. Sea stars typically have five arms, but they may have more (Figure 23-1D), and there are as many ambulacral areas as there are arms. An ambulacral groove is found along the middle of each ambulacral area, and the groove is bordered by rows of tube feet (podia) (Figure 23-2). These in turn are usually protected by movable spines. A large radial nerve can be seen in the center of each ambulacral groove (Figure 23-3C), between the rows of tube feet. The nerve is very superficially located, covered only by thin epidermis. Under the nerve is an extension of the coelom and the radial canal of the water-vascular system, all of which are external to the underlying ossicles (Figure 23-3C). In all other classes of living echinoderms except crinoids, these structures are covered by ossicles or other dermal tissue; thus ambulacral grooves in asteroids and crinoids are said to be open, and those of the other groups are closed.
 |
| Figure 23-2 External anatomy of asteroid. A, Aboral view. B, Oral view. |
 |
| Figure 23-3 A, Internal anatomy of a sea star. B, Water-vascular system. Podia penetrate between ossicles. (Polian vesicles are not present in Asterias.) C, Cross section of arm at level of gonads, illustrating open ambulacral grooves. |
The aboral surface is usually rough and spiny, although spines of many species are flattened, so that the surface appears smooth (Figure 23-1C). Around the bases of spines are groups of minute, pincerlike pedicellariae, bearing tiny jaws manipulated by muscles (Figure 23-4). These jaws help keep the body surface free of debris, protect the papulae, and sometimes aid in food capture. Papulae (dermal branchiae or skin gills) are soft delicate projections of the coelomic cavity, covered only with epidermis and lined internally with peritoneum; they extend out through spaces between ossicles and are concerned with respiration (Figures 23-3C, and 23-4F). Also on the aboral side are the inconspicuous anus and the circular madreporite (Figure 23-2A), a calcareous sieve leading to the water-vascular system.
 |
|
 |
Figure 23-4 Pedicellariae of sea stars and sea urchins. A, Forceps-type pedicellaria of Asterias. B, and C, Scissors-type pedicellariae of Asterias; size relative to spine is shown in B. D, Tridactyl pedicellaria of Strongylocentrotus. E, Globiferous pedicellaria of Strongylocentrotus. F, Close-up view of the aboral surface of the sea star Pycnopodia helianthoides. Note the large pedicellariae, as well as groups of small pedicellariae around the spines. Many thinwalled papulae can be seen. |
Endoskeleton
Beneath the epidermis of sea stars is a mesodermal endoskeleton of small calcareous plates, or ossicles, bound together with connective tissue. From these ossicles project the spines and tubercles that make up the spiny surface. Ossicles are penetrated by a meshwork of spaces, usually filled with fibers and dermal cells. This internal meshwork structure is described as stereom and is unique to echinoderms.
Muscles in the body wall move the rays and can partially close the ambulacral grooves by drawing their margins together.
Coelom, Excretion, and Respiration
The coelomic compartments of larval echinoderms give rise to several structures in adults, one of which is a spacious body coelom filled with fluid. The fluid contains amebocytes (coelomocytes), bathes the internal organs, and projects into the papulae. The ciliated peritoneal lining of the coelom circulates the fluid around the body cavity and into the papulae. Exchange of respiratory gases and excretion of nitrogenous waste, principally ammonia, take place by diffusion through the thin walls of papulae and tube feet. Some wastes may be picked up by coelomocytes, which migrate through the epithelium of the papulae or tube feet to the exterior, or the tips of papulae containing waste-laden coelomocytes may pinch off.
Water-Vascular System
The water-vascular system is another coelomic compartment and is unique to echinoderms. Showing exploitation of hydraulic mechanisms to a greater degree than in any other animal group, it is a system of canals and specialized tube feet that, together with the dermal ossicles, has determined the evolutionary potential and limitations of this phylum. In sea stars the primary functions of the water-vascular system are locomotion and food gathering, in addition to respiration and excretion.
Structurally, the water-vascular system opens to the outside through small pores in the madreporite. The madreporite of asteroids is on the aboral surface (Figure 23-2A) and leads into the stone canal, which descends toward the ring canal around the mouth (Figure 23-3B). Radial canals diverge from the ring canal, one into the ambulacral groove of each ray. Also attached to the ring canal are four or five pairs of folded, pouchlike Tiedemann’s bodies and from one to five polian vesicles (polian vesicles are absent in some sea stars, such as Asterias). Tiedemann’s bodies may produce coelomocytes, and polian vesicles are apparently for fluid storage.
A series of small lateral canals, each with a one-way valve, connects the radial canal to the cylindrical podia, or tube feet, along the sides of the ambulacral groove in each ray. Each podium is a hollow, muscular tube, the inner end of which is a muscular sac, or ampulla, that lies within the body coelom (Figure 23-3A and C), and the outer end of which usually bears a sucker. Some species lack suckers. Podia pass to the outside between ossicles in the ambulacral groove.
The water-vascular system operates hydraulically and is an effective locomotor mechanism. The valves in the lateral canals prevent backflow of fluid into the radial canals. Each tube foot has in its walls connective tissue that maintains the cylinder at a relatively constant diameter. Contraction of muscles in the ampulla forces fluid into the podium, extending it. Conversely, contraction of the longitudinal muscles in the tube foot retracts the podium, forcing fluid back into the ampulla. Contraction of muscles in one side of the podium bends the organ toward that side. Small muscles at the end of the tube foot can raise the middle of the disclike end, creating suction when the end is applied to a firm substrate. It has been estimated that by combining mucous adhesion with suction, a single podium can exert a pull equal to 25 to 30 g. Coordinated action of all or many of the tube feet is sufficient to draw the animal up a vertical surface or over rocks. The ability to move while firmly adhering to the substrate is a clear advantage to an animal living in a sometimes wave-churned environment.
On a soft surface, such as muck or sand, suckers are ineffective (numerous sand-dwelling species have no suckers), so the tube feet are employed as legs. Locomotion becomes mainly a stepping process. Most sea stars can move only a few centimeters per minute, but some very active ones can move 75 to 100 cm per minute; for example, Pycnopodia (Gr. pyknos, compact, dense, + pous, podos, foot) (Figure 23-5B). When inverted, a sea star bends its rays until some of the tubes reach the substratum and attach as an anchor; then it slowly rolls over.
Tube feet are innervated by the central nervous system (ectoneural and hyponeural systems, see following text). Nervous coordination enables tube feet to move in a single direction, although not in unison, so that the sea star may progress. If the radial nerve in an arm is cut, podia in that arm lose coordination, although they can still function. If the circumoral nerve ring is cut, podia in all arms become uncoordinated, and movement ceases.
 |
| Figure 23-5 A, Orthasterias koehleri eating a clam. B, This Pycnopodia helianthoides has been overturned while eating a large sea urchin Strongylocentrotus franciscanus. This sea star has 20 to 24 arms and can range up to 1 m in diameter (arm tip to arm tip). |
 |
| Figure 23-6 Crown-of-thorns star Acanthaster planci feeding on coral. Puncture wounds from its spines are painful; the spines are equipped with poison glands. |
The mouth on the oral side leads through a short esophagus to a large stomach in the central disc. The lower (cardiac) part of the stomach can be everted through the mouth during feeding (Figure 23-2B), and excessive eversion is prevented by gastric ligaments. The upper (pyloric) part is smaller and connects by ducts to a pair of large pyloric ceca (digestive glands) in each arm (Figure 23-3A). Digestion is mostly extracellular, although some intracellular digestion may occur in the ceca. A short intestine leads aborally from the pyloric stomach, and there are usually a few small, saclike intestinal ceca (Figure 23-3A). The anus is inconspicuous, and some sea stars lack an intestine and anus.
Many sea stars are carnivorous and feed on molluscs, crustaceans, polychaetes, echinoderms, other invertebrates, and sometimes small fish. Sea stars consume a wide range of food items, but many show particular preferences (Figures 23-5 and 23-6). Some select brittle stars, sea urchins, or sand dollars, swallowing them whole and later regurgitating undigestible ossicles and spines (Figure 23-5B). Some attack other sea stars, and if they are small compared with their prey, they may attack and begin eating at the end of one arm.
Some asteroids feed heavily on molluscs (Figure 23-5A), and Asterias is a significant predator on commercially important clams and oysters. When feeding on a bivalve, a sea star will hump over its prey, attaching its podia to the valves, and then exert a steady pull, using its feet in relays. A force of some 1300 g can thus be exerted. In half an hour or so the adductor muscles of the bivalve fatigue and relax. With a very small gap available, the star inserts its soft everted stomach into the space between the valves and wraps it around the soft parts of the shellfish. After feeding, the sea star draws its stomach inward by contraction of the stomach muscles and relaxation of body-wall muscles.
Some sea stars feed on small particles, either entirely or in addition to carnivorous feeding. Plankton and other organic particles coming in contact with the animal’s surface are carried by the epidermal cilia to the ambulacral grooves and then to the mouth.
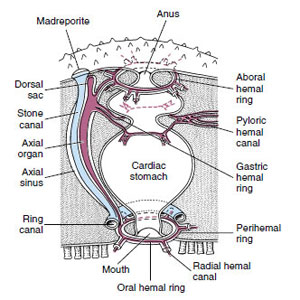 |
| Figure 23-7 Hemal system of asteroids. The main perihemal channel is the thin-walled axial sinus, which encloses both the axial organ and the stone canal. Other features of the hemal system are shown. |
Hemal System
The so-called hemal system is not very well developed in asteroids, and its function in all echinoderms is unclear. The hemal system has little or nothing to do with circulation of body fluids. It is a system of tissue strands enclosing unlined sinuses and is itself enclosed in another coelomic compartment, the perihemal channels (Figure 23-7). The hemal system may be useful in distributing digested products, but its specific functions are not really known.
Nervous System
The nervous system consists of three units at different levels in the disc and arms. Chief of these systems is the oral (ectoneural) system composed of a nerve ring around the mouth and a main radial nerve into each arm. It appears to coordinate the tube feet. A deep (hyponeural) system lies aboral to the oral system, and an aboral system consists of a ring around the anus and radial nerves along the roof of each ray. An epidermal nerve plexus or nerve net freely connects these systems with the body wall and related structures. The epidermal plexus coordinates responses of the dermal branchiae to tactile stimulation—the only instance known in echinoderms in which coordination occurs through a nerve net.
Sense organs are not well developed. Tactile organs and other sensory cells are scattered over the surface, and an ocellus is at the tip of each arm. Their reactions are mainly to touch, temperature, chemicals, and differences in light intensity. Sea stars are usually more active at night.
Reproductive System, Regeneration, and Autotomy
Most sea stars have separate sexes. A pair of gonads lies in each interradial space (Figure 23-3A). Fertilization is external and occurs in early summer when eggs and sperm are shed into the water. A secretion from neurosecretory cells located on the radial nerves stimulates maturation and shedding of asteroid eggs.
 |
| Figure 23-8 Pacific sea star Echinaster luzonicus can reproduce itself by splitting across the disc, then regenerating missing arms. The one shown here has evidently regenerated six arms from the longer one at top left. |
Echinoderms can regenerate lost parts. Sea star arms can regenerate readily, even if all are lost. Sea stars also have the power of autotomy and can cast off an injured arm near the base. Regeneration of a new arm may take several months.
Some species can regenerate a complete new sea star (Figure 23-8) from a detached arm that contains a part (about one-fifth) of the central disc. In former times fishermen used to dispatch sea stars they collected from their oyster beds by chopping them in half with a hatchet—a worse than futile activity. Some sea stars reproduce asexually under normal conditions by cleaving the central disc, each part regenerating the rest of the disc and missing arms.
Development
In some species the liberated eggs are brooded, either under the oral side of the animal or in specialized aboral structures, and development is direct, but in most species embryonating eggs are free in the water and hatch to freeswimming larvae.
Early embryogenesis shows a typical primitive deuterostome pattern (see Figures 8-7A and 8-11A). Gastrulation is by invagination, and the anterior end of the archenteron pinches off to become the coelomic cavity, which expands in a U shape to fill the blastocoel. Each of the legs of the U, at the posterior, constricts to become a separate vesicle, and these eventually give rise to the main coelomic compartments of the body (metacoels, called somatocoels in echinoderms). The anterior portion of the U undergoes subdivision to form the protocoels and mesocoels (called axocoels and hydrocoels in echinoderms) (Figure 23-9). The left hydrocoel will become the water-vascular system, and the left axocoel will give rise to the stone canal and perihemal channels. The right axocoel and hydrocoel will disappear. The free-swimming larva has cilia arranged in bands and is called a bipinnaria (Figure 23-10A). These ciliated tracts become extended into larval arms. Soon the larva grows three adhesive arms and a sucker at its anterior end and is then called a brachiolaria. At that time it attaches to the substratum, forms a temporary attachment stalk, and undergoes metamorphosis.
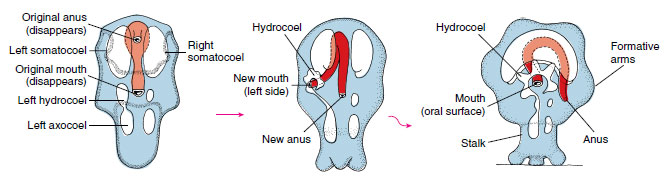 |
| Figure 23-9 Asteroid metamorphosis. The left somatocoel becomes the oral coelom, and the right somatocoel becomes the aboral coelom. The left hydrocoel becomes the water-vascular system and the left axocoel the stone canal and perihemal channels. The right axocoel and hydrocoel are lost. |
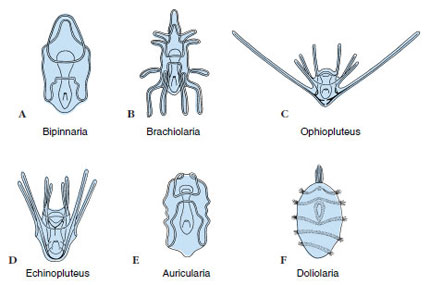 |
| Figure 23-10 Larvae of echinoderms. A, Bipinnaria of asteroids. B, Brachiolaria of asteroids. C, Ophiopluteus of ophiuroids. D, Echinopluteus of echinoids. E, Auricularia of holothuroids. F, Doliolaria of crinoids. |
Metamorphosis involves a dramatic reorganization of a bilateral larva into a radial juvenile. The anteroposterior axis of the larva is lost, and what was the left side becomes the oral surface, and the larval right side becomes the aboral surface (Figure 23-9). Correspondingly, the larval mouth and anus disappear, and a new mouth and anus form on what were originally the left and right sides, respectively. The portion of the anterior coelomic compartment from the left side expands to form the ring canal of the watervascular system around the mouth, and then it grows branches to form the radial canals. As the short, stubby arms and the first podia appear, the animal detaches from its stalk and begins life as a young sea star.
Support our developers




