Phylogeny and Adaptive Radiation
Phylogeny and
Adaptive Radiation
Phylogeny
Despite the existence of an extensive fossil record, there have been numerous contesting hypotheses on echinoderm phylogeny. Based on the embryological evidence of the bilateral larvae, there can be little doubt that their ancestors were bilateral and that their coelom had three pairs of spaces (trimeric). Some investigators have held that radial symmetry arose in a free-moving echinoderm ancestor and that sessile groups were derived several times independently from the freemoving ancestors. However, this view does not account for the adaptive significance of radial symmetry as an adaptation for a sessile existence. The more traditional view is that the first echinoderms were sessile, became radial as an adaptation to that existence, and then gave rise to the freemoving groups. Figure 23-29 is consistent with this hypothesis. It views evolution of endoskeletal plates with stereom structure and of external ciliary grooves for feeding as early echinoderm (or pre-echinoderm) developments. The extinct carpoids (Figures 23-28A, 23-29) had stereom ossicles but were not radially symmetrical, and the status of their water-vascular system, if any, is uncertain. Some investigators regard carpoids as a separate subphylum of echinoderms (Homalozoa) and consider them closer to chordates (Calcichordata). The fossil helicoplacoids (Figures 23-28B, 23-29) show evidence of three, true ambulacral grooves, and their mouth was on the side of the body.
Attachment to the substratum by the aboral surface would have led to radial symmetry and the origin of the Pelmatozoa. Both Cystoidea (extinct) and Crinoidea primitively were attached to the substratum by an aboral stalk. An ancestor that became freemoving and applied its oral surface to the substratum would have given rise to Eleutherozoa. Phylogeny within Eleutherozoa is controversial. Most investigators agree that echinoids and holothuroids are related and form a clade, but opinions diverge on the relationship of ophiuroids and asteroids. Figure 23-29 illustrates the view that the ophiuroids arose after the closure of ambulacral grooves, but this scheme treats evolution of five ambulacral rays (arms) in ophiuroids and asteroids as independently evolved. Alternatively, if ophiuroids and asteroids are a single clade, then closed ambulacral grooves must have evolved separately in ophiuroids and in the common ancestor of echinoids and holothuroids.
Data on the Concentricycloidea are insufficient to place this group on a cladogram, although they are tentatively placed in Eleutherozoa.
Adaptive Radiation
Radiation of echinoderms has been determined by limitations and potentials inherent in their most important characters: radial symmetry, watervascular system, and dermal endoskeleton. If their ancestors had a brain and specialized sense organs, these were lost in the adoption of radial symmetry. Thus it is not surprising that there are large numbers of creeping, benthic forms with filter-feeding, deposit-feeding, scavenging, and herbivorous habits, comparatively few predators, and very rare pelagic forms. In this light the relative success of asteroids as predators is impressive and probably attributable to the extent to which they have exploited the hydraulic mechanism of their tube feet.
The basic body plan of echinoderms has severely limited their evolutionary opportunities to become parasites. Indeed, the most mobile of echinoderms, the ophiuroids, which are also the ones most able to insert their bodies into small spaces, are the only group with significant numbers of commensal species.
Classification of Phylum Echinodermata
There are about 6,000 living and 20,000 extinct or fossil species of Echinodermata. The traditional classification placed all free-moving forms that were oriented with oral side down in subphylum Eleutherozoa, containing most living species. The other subphylum, Pelmatozoa, contained mostly forms with stems and oral side up; most extinct classes and living Crinoidea belong to this group. Although alternative schemes have strong supporters, cladistic analysis provides evidence that the two traditional subphyla are monophyletic.* The following includes only groups with living members.
Subphylum Pelmatozoa (pel-ma´tozo ´a) (Gr. pelmatos; a stalk, + zoon, animal). Body in form of cup or calyx, borne on aboral stalk during part or all of life; oral surface directed upward; open ambulacral grooves; madreporite absent; both mouth and anus on oral surface; several fossil classes plus living Crinoidea.
Phylogeny
Despite the existence of an extensive fossil record, there have been numerous contesting hypotheses on echinoderm phylogeny. Based on the embryological evidence of the bilateral larvae, there can be little doubt that their ancestors were bilateral and that their coelom had three pairs of spaces (trimeric). Some investigators have held that radial symmetry arose in a free-moving echinoderm ancestor and that sessile groups were derived several times independently from the freemoving ancestors. However, this view does not account for the adaptive significance of radial symmetry as an adaptation for a sessile existence. The more traditional view is that the first echinoderms were sessile, became radial as an adaptation to that existence, and then gave rise to the freemoving groups. Figure 23-29 is consistent with this hypothesis. It views evolution of endoskeletal plates with stereom structure and of external ciliary grooves for feeding as early echinoderm (or pre-echinoderm) developments. The extinct carpoids (Figures 23-28A, 23-29) had stereom ossicles but were not radially symmetrical, and the status of their water-vascular system, if any, is uncertain. Some investigators regard carpoids as a separate subphylum of echinoderms (Homalozoa) and consider them closer to chordates (Calcichordata). The fossil helicoplacoids (Figures 23-28B, 23-29) show evidence of three, true ambulacral grooves, and their mouth was on the side of the body.
Attachment to the substratum by the aboral surface would have led to radial symmetry and the origin of the Pelmatozoa. Both Cystoidea (extinct) and Crinoidea primitively were attached to the substratum by an aboral stalk. An ancestor that became freemoving and applied its oral surface to the substratum would have given rise to Eleutherozoa. Phylogeny within Eleutherozoa is controversial. Most investigators agree that echinoids and holothuroids are related and form a clade, but opinions diverge on the relationship of ophiuroids and asteroids. Figure 23-29 illustrates the view that the ophiuroids arose after the closure of ambulacral grooves, but this scheme treats evolution of five ambulacral rays (arms) in ophiuroids and asteroids as independently evolved. Alternatively, if ophiuroids and asteroids are a single clade, then closed ambulacral grooves must have evolved separately in ophiuroids and in the common ancestor of echinoids and holothuroids.
Data on the Concentricycloidea are insufficient to place this group on a cladogram, although they are tentatively placed in Eleutherozoa.
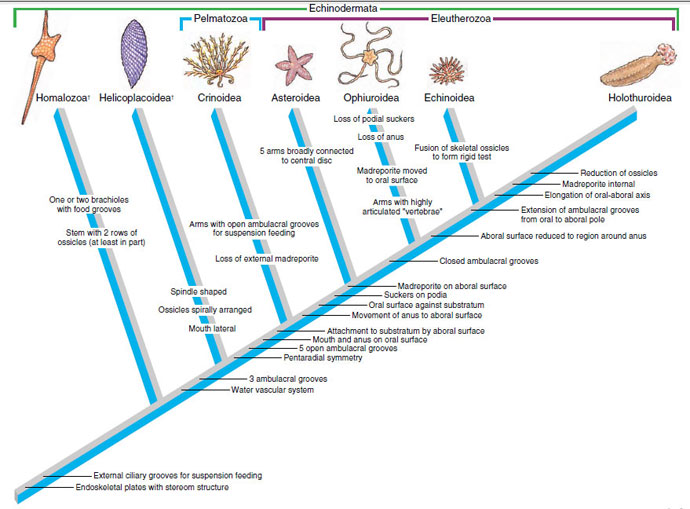 |
| Figure 23-29 Cladogram showing hypothetical relationships among echinoderm groups. The extinct Homalozoa (carpoids), which were not radial in symmetry but had stereom endoskeletal plates, represent an early split from echinoderms. An intermediate form is represented by the extinct helicoplacoids, which had three ambulacral grooves that wound around their bodies in a spiral fashion. Helicoplacoids are the sister group of modern echinoderms. Evolution of pentaradial symmetry was an adaptation to sessile existence and is a synapomorphy of modern echinoderms. The scheme depicted here views ophiuroids as having arisen separately from asteroids, after the evolution of closed ambulacral grooves, and the possession of five arms would thus have been of separate origin. Alternatively, if Asteroidea and Ophiuroidea form a monophyletic group, with five arms being synapomorphic, then closed ambulacral grooves in the ophiuroids would have evolved separately from that character in echinoids and holothuroids |
Adaptive Radiation
Radiation of echinoderms has been determined by limitations and potentials inherent in their most important characters: radial symmetry, watervascular system, and dermal endoskeleton. If their ancestors had a brain and specialized sense organs, these were lost in the adoption of radial symmetry. Thus it is not surprising that there are large numbers of creeping, benthic forms with filter-feeding, deposit-feeding, scavenging, and herbivorous habits, comparatively few predators, and very rare pelagic forms. In this light the relative success of asteroids as predators is impressive and probably attributable to the extent to which they have exploited the hydraulic mechanism of their tube feet.
The basic body plan of echinoderms has severely limited their evolutionary opportunities to become parasites. Indeed, the most mobile of echinoderms, the ophiuroids, which are also the ones most able to insert their bodies into small spaces, are the only group with significant numbers of commensal species.
Classification of Phylum Echinodermata
There are about 6,000 living and 20,000 extinct or fossil species of Echinodermata. The traditional classification placed all free-moving forms that were oriented with oral side down in subphylum Eleutherozoa, containing most living species. The other subphylum, Pelmatozoa, contained mostly forms with stems and oral side up; most extinct classes and living Crinoidea belong to this group. Although alternative schemes have strong supporters, cladistic analysis provides evidence that the two traditional subphyla are monophyletic.* The following includes only groups with living members.
Subphylum Pelmatozoa (pel-ma´tozo ´a) (Gr. pelmatos; a stalk, + zoon, animal). Body in form of cup or calyx, borne on aboral stalk during part or all of life; oral surface directed upward; open ambulacral grooves; madreporite absent; both mouth and anus on oral surface; several fossil classes plus living Crinoidea.
-
Class Crinoidea (krin-oi´de-a) (Gr. krinon, lily; + eidos, form; + ea,
characterized by): sea lilies and feather stars. Five arms branching at
base and bearing pinnules; ciliated
ambulacral grooves on oral surface
with tentacle-like tube feet for food
gathering; spines, madreporite, and
pedicellariae absent. Examples: Antedon,
Comantheria (Figure 23-26).
-
Class Concentricycloidea (konsen
´tri-sy-kloy´de-a) (L. cum, together,
+ centrum, center [having a common
center], + Gr. kyklos, circle, + eidos, form, + ea, characterized by): sea daisies. Disc-shaped body, with
marginal spines but no arms; concentrically
arranged skeletal plates; ring
of suckerless podia near body margin;
hydropore present; gut present or
absent, no anus. Example: Xyloplax (Figure 23-27).
Class Asteroidea (as´ter-oy´de-a) (Gr. aster, star, + eidos, form, + ea, characterized by): sea stars (starfish). Star-shaped, with arms not sharply marked off from central disc; ambulacral grooves open, with tube feet on oral side; tube feet often with suckers; anus and madreporite aboral; pedicellariae present. Examples: Asterias, Pisaster.
Class Ophiuroidea (o´fe-u-roy´de-a) (Gr. ophis, snake, + oura, tail, + eidos, form, + ea, characterized by): brittle stars and basket stars. Star shaped, with arms sharply marked off from central disc; ambulacral grooves closed, covered by ossicles; tube feet without suckers and not used for locomotion; pedicellariae absent; anus absent. Examples: Ophiura (Figure 23-11A), Gorgonocephalus (Figure 23-14B).
Class Echinoidea (ek´i-noy´de-a) (Gr. echinos, sea urchin, hedgehog, + eidos, form, + ea, characterized by): sea urchins, sea biscuits, and sand dollars. More or less globular or discshaped, with no arms; compact skeleton or test with closely fitting plates; movable spines; ambulacral grooves closed; tube feet with suckers; pedicellariae present. Examples: Arbacia, Strongylocentrotus (Figure 23-15), Lytechinus, Mellita.
Class Holothuroidea (hol´o-thuroy ´de-a) (Gr. holothourion, sea cucumber, + eidos, form, + ea, characterized by): sea cucumbers. Cucumber-shaped, with no arms; spines absent; microscopic ossicles embedded in thick muscular wall; anus present; ambulacral grooves closed; tube feet with suckers; circumoral tentacles (modified tube feet); pedicellariae absent; madreporite internal. Examples: Sclerodactyla, Parastichopus, Cucumaria (Figure 23-21C).
Support our developers




