Lungs
Lungs
Gills are unsuitable for life in air because, when removed from the buoying water medium, gill filaments collapse, dry, and stick together; a fish out of water rapidly asphyxiates despite the abundance of oxygen around it. Consequently most airbreathing vertebrates possess lungs, highly vascularized internal cavities. Lungs of a sort are found in certain invertebrates (pulmonate snails, scorpions, some spiders, some small crustaceans), but these structures cannot be very efficiently ventilated.
Lungs that can be ventilated by muscle movements to produce a rhythmic exchange of air are characteristic of terrestrial vertebrates. Most rudimentary of vertebrate lungs are those of lungfishes (Dipneusti), which use them to supplement, or even replace, gill respiration during periods of drought. Although of simple construction, a lungfish lung is supplied with a capillary network in its largely unfurrowed walls, a tubelike connection to the pharynx, and a primitive ventilating system for moving air in and out of the lung.
Amphibian lungs vary from simple, smooth-walled, baglike lungs of some salamanders to the subdivided lungs of frogs and toads (Figure 33-21). The total surface available for gas exchange is much increased in lungs of reptiles which are subdivided into numerous interconnecting air sacs. Most elaborate of all are mammalian lungs complexes of millions of small sacs, called alveoli (Figure 33-22), each veiled by a rich vascular network. Human lungs have a total surface area of from 50 to 90 m2— 50 times the area of the skin surface— and contain 1000 km of capillaries. A large surface area is essential for the high oxygen uptake required to support the elevated metabolic rate of endothermic mammals.
A disadvantage of lungs is that gas is exchanged between blood and air only in the alveoli, located at the ends of a branching tree of air tubes (trachea, bronchi, and bronchioles [Figure 33-22]). Unlike the efficient one-way flow of water across fish gills, air must enter and exit a lung through the same channel. After exhalation, the air tubes are filled with “used” air from the alveoli which, during the following inhalation, is pulled back into the lungs. The volume of air in a lung’s passageways is called “dead space.” This air shuttles back and forth with each breath, adding to the difficulty of properly ventilating lungs. In fact, lung ventilation in humans is so inefficient that in normal breathing only approximately one-sixth of the air in the lungs is replenished with each inspiration. Even after forced expiration, 20% to 35% of the air remains in the lungs.
In birds, lung efficiency is improved vastly by adding an extensive system of air sacs that serve as air reservoirs during ventilation. On inspiration, some 75% of incoming air bypasses the lungs to enter the air sacs (gas exchange does not occur here). At expiration some of this fresh air passes directly through the lung passages and eventually into one-cell thick air capillaries where gas exchange occurs. Thus air capillaries receive nearly fresh air during both inspiration and expiration. The beautifully designed bird lung is a result of selective pressures during evolution of flight with its high metabolic demands.
Amphibians employ a positive pressure action to force air into their lungs, unlike most reptiles, birds, and mammals, which ventilate their lungs by negative pressure, in which air is pulled into the lungs by expansion of the thoracic cavity. Frogs ventilate the lungs by first drawing air into the mouth through the external nares (nostrils). Then, closing the nares and raising the floor of the mouth, they drive air into the lungs (Figure 33-23). Much of the time, however, frogs rhythmically ventilate only the mouth cavity, a well-vascularized respiratory surface that supplements pulmonary respiration.
Gills are unsuitable for life in air because, when removed from the buoying water medium, gill filaments collapse, dry, and stick together; a fish out of water rapidly asphyxiates despite the abundance of oxygen around it. Consequently most airbreathing vertebrates possess lungs, highly vascularized internal cavities. Lungs of a sort are found in certain invertebrates (pulmonate snails, scorpions, some spiders, some small crustaceans), but these structures cannot be very efficiently ventilated.
Lungs that can be ventilated by muscle movements to produce a rhythmic exchange of air are characteristic of terrestrial vertebrates. Most rudimentary of vertebrate lungs are those of lungfishes (Dipneusti), which use them to supplement, or even replace, gill respiration during periods of drought. Although of simple construction, a lungfish lung is supplied with a capillary network in its largely unfurrowed walls, a tubelike connection to the pharynx, and a primitive ventilating system for moving air in and out of the lung.
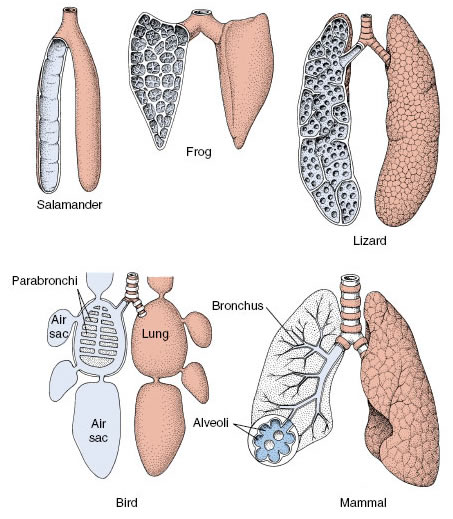 |
| Figure 33-21 Internal structures of lungs among vertebrate groups. Generally, the evolutionary trend has been from simple sacs with little exchange surface between blood and air spaces to complex, lobulated structures, each with complex divisions and extensive exchange surfaces. |
Amphibian lungs vary from simple, smooth-walled, baglike lungs of some salamanders to the subdivided lungs of frogs and toads (Figure 33-21). The total surface available for gas exchange is much increased in lungs of reptiles which are subdivided into numerous interconnecting air sacs. Most elaborate of all are mammalian lungs complexes of millions of small sacs, called alveoli (Figure 33-22), each veiled by a rich vascular network. Human lungs have a total surface area of from 50 to 90 m2— 50 times the area of the skin surface— and contain 1000 km of capillaries. A large surface area is essential for the high oxygen uptake required to support the elevated metabolic rate of endothermic mammals.
A disadvantage of lungs is that gas is exchanged between blood and air only in the alveoli, located at the ends of a branching tree of air tubes (trachea, bronchi, and bronchioles [Figure 33-22]). Unlike the efficient one-way flow of water across fish gills, air must enter and exit a lung through the same channel. After exhalation, the air tubes are filled with “used” air from the alveoli which, during the following inhalation, is pulled back into the lungs. The volume of air in a lung’s passageways is called “dead space.” This air shuttles back and forth with each breath, adding to the difficulty of properly ventilating lungs. In fact, lung ventilation in humans is so inefficient that in normal breathing only approximately one-sixth of the air in the lungs is replenished with each inspiration. Even after forced expiration, 20% to 35% of the air remains in the lungs.
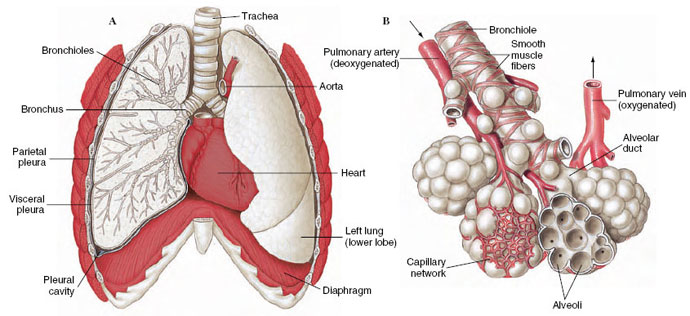 |
| Figure 33-22 A, Lungs of human with right lung shown in section. B, Terminal portion of bronchiole showing air sacs with their blood supply. Arrows show direction of blood flow. |
In birds, lung efficiency is improved vastly by adding an extensive system of air sacs that serve as air reservoirs during ventilation. On inspiration, some 75% of incoming air bypasses the lungs to enter the air sacs (gas exchange does not occur here). At expiration some of this fresh air passes directly through the lung passages and eventually into one-cell thick air capillaries where gas exchange occurs. Thus air capillaries receive nearly fresh air during both inspiration and expiration. The beautifully designed bird lung is a result of selective pressures during evolution of flight with its high metabolic demands.
Amphibians employ a positive pressure action to force air into their lungs, unlike most reptiles, birds, and mammals, which ventilate their lungs by negative pressure, in which air is pulled into the lungs by expansion of the thoracic cavity. Frogs ventilate the lungs by first drawing air into the mouth through the external nares (nostrils). Then, closing the nares and raising the floor of the mouth, they drive air into the lungs (Figure 33-23). Much of the time, however, frogs rhythmically ventilate only the mouth cavity, a well-vascularized respiratory surface that supplements pulmonary respiration.
Support our developers




