Phylogeny and Adaptive Radiation
Phylogeny and
Adaptive Radiation
Insect fossils, although not abundant, have been found in numbers sufficient to give a general idea of the evolutionary history of insects. Although several groups of marine arthropods, such as trilobites, crustaceans, and xiphosurans, were present in the Cambrian period, the first terrestrial arthropods—scorpions and millipedes—did not appear until the Silurian period. The first insects, which were wingless, date from the Devonian period. By the Carboniferous period, several orders of winged insects, most of which are now extinct, had appeared.
Opinions of arthropodologists on relationships among the animals composing Uniramia are diverse. Many scientists are disinclined to use the name at all because it was originally proposed to include Onychophora. We are using the term as a convenience, but we suggest it should be accepted with suitable caution. Figure 20-38 is only one hypothesis. Some scientists believe that Myriapoda is paraphyletic and that the Diplopoda/Pauropoda clade is the sister group of insects. Some molecular data and data based on enervation of the eyes suggest a more remote relationship of insects and myriapods and that insects may have arisen within subphylum Crustacea. Some sequence analyses suggest that myriapods share a more recent common ancestor with chelicerates than with insects.
Whether or not some or all myriapods are closely related to insects, it seems likely that the ancestral insect had a head and trunk of many similar somites, a primitive character retained by myriapods. Evolution of insects involved specialization of the first three postcephalic somites to become locomotor somites (thorax) and a loss or reduction of appendages on the rest of the body (abdomen). The wingless apterygotes have traditionally been regarded as having the most primitive characteristics, but subclass Apterygota is apparently paraphyletic (Figure 20-39). Three apterygote orders (Diplura, Collembola, Protura) have their mandibles and first maxillae located deeply in pouches in the head, a condition known as endognathy. They share other primitive and derived characters, and there are many similarities between endognathous insects and myriapods. All other insects are ectognathous, including the wingless order Thysanura. Ectognathous insects do not have their mandibles and maxillae in pouches, and they share other synapomorphies. Endognathous and ectognathous insects form sister groups, and Thysanura diverged from the common ancestor of ectognathous insects before the advent of flight,* which unites the remaining ectognathous orders. The evolutionary origin of insect wings has long been a puzzle. The adaptive value of wings for flight is clear, but such structures do not spring into existence fully developed. They must have evolved from earlier structures that would have been too small to support flight, but what possible value to their owners were “wings” too small to fly? One plausible suggestion is that proto-wings were used by their possessors for skimming across the surface of water, much as present-day stoneflies use their wings.
The ancestral winged insect gave rise to three lines, which differed in their ability to flex their wings. Two of these (Odonata and Ephemeroptera) have outspread wings or hold their wings vertically over the abdomen. The other line has wings that can fold back horizontally over the abdomen. It branched into three groups, all of which were present by the Permian period. One group with hemimetabolous metamorphosis, chewing mouthparts, and cerci includes Orthoptera, Dermaptera, Isoptera, and Embioptera; another group with hemimetabolous metamorphosis and a tendency toward sucking mouthparts includes Thysanoptera, Hemiptera, and Homoptera and perhaps also Psocoptera, Zoraptera, Mallophaga, and Anoplura, although there is some disagreement among authorities about the last group. Insects with holometabolous metamorphosis have the most specialized life history, and these apparently form a clade.
The adaptive properties of insects have been stressed throughout this section. The directions and ranges of their adaptive radiation, both structurally and physiologically, have been amazingly varied. Whether it be in the area of habitat, feeding adaptations, means of locomotion, reproduction, or general mode of living, the adaptive achievements of insects are truly remarkable.
Insect fossils, although not abundant, have been found in numbers sufficient to give a general idea of the evolutionary history of insects. Although several groups of marine arthropods, such as trilobites, crustaceans, and xiphosurans, were present in the Cambrian period, the first terrestrial arthropods—scorpions and millipedes—did not appear until the Silurian period. The first insects, which were wingless, date from the Devonian period. By the Carboniferous period, several orders of winged insects, most of which are now extinct, had appeared.
Opinions of arthropodologists on relationships among the animals composing Uniramia are diverse. Many scientists are disinclined to use the name at all because it was originally proposed to include Onychophora. We are using the term as a convenience, but we suggest it should be accepted with suitable caution. Figure 20-38 is only one hypothesis. Some scientists believe that Myriapoda is paraphyletic and that the Diplopoda/Pauropoda clade is the sister group of insects. Some molecular data and data based on enervation of the eyes suggest a more remote relationship of insects and myriapods and that insects may have arisen within subphylum Crustacea. Some sequence analyses suggest that myriapods share a more recent common ancestor with chelicerates than with insects.
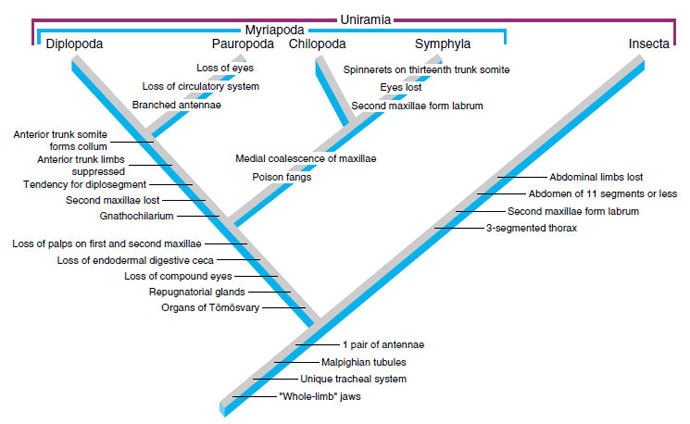 |
| Figure 20-38 Cladogram showing hypothetical relationships of uniramians. Here the myriapods and insects are sister groups; therefore Diplopoda, Pauropoda, Chilopoda, and Symphyla would become subclasses under the class Myriapoda. Another hypothesis would make the Diplopoda and Pauropoda class the sister group of insects. Organs of Tömösvary are unique sensory organs opening at the bases of the antennae, and repugnatorial glands, located on certain somites or legs, secrete an obnoxious substance for defense. The gnathochilarium is formed in diplopods and pauropods by fusion of the first maxillae, and the collum is the collarlike tergite of the first trunk segment. Formation of a labrum from the second maxillae has been sometimes considered evidence of sister-group relationship of symphylans and insects; it is viewed here as convergence. Outgroups for this cladogram would be non-uniramian arthropod lineages. |
Whether or not some or all myriapods are closely related to insects, it seems likely that the ancestral insect had a head and trunk of many similar somites, a primitive character retained by myriapods. Evolution of insects involved specialization of the first three postcephalic somites to become locomotor somites (thorax) and a loss or reduction of appendages on the rest of the body (abdomen). The wingless apterygotes have traditionally been regarded as having the most primitive characteristics, but subclass Apterygota is apparently paraphyletic (Figure 20-39). Three apterygote orders (Diplura, Collembola, Protura) have their mandibles and first maxillae located deeply in pouches in the head, a condition known as endognathy. They share other primitive and derived characters, and there are many similarities between endognathous insects and myriapods. All other insects are ectognathous, including the wingless order Thysanura. Ectognathous insects do not have their mandibles and maxillae in pouches, and they share other synapomorphies. Endognathous and ectognathous insects form sister groups, and Thysanura diverged from the common ancestor of ectognathous insects before the advent of flight,* which unites the remaining ectognathous orders. The evolutionary origin of insect wings has long been a puzzle. The adaptive value of wings for flight is clear, but such structures do not spring into existence fully developed. They must have evolved from earlier structures that would have been too small to support flight, but what possible value to their owners were “wings” too small to fly? One plausible suggestion is that proto-wings were used by their possessors for skimming across the surface of water, much as present-day stoneflies use their wings.
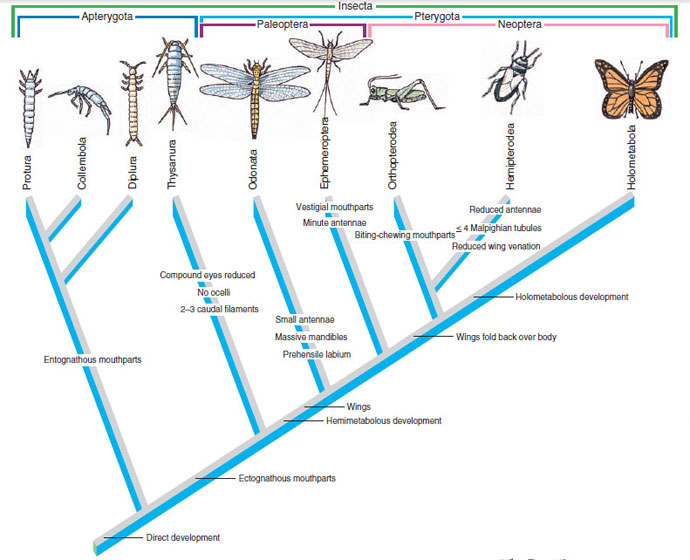 |
| Figure 20-39 Cladogram showing hypothetical relationships among insects. Many synapomorphies have been omitted. Orders Protura, Collembola, and Diplura are entognathous, and together with the ectognathous order Thysanura form the traditional (but paraphyletic) group Apterygota. These orders originated before the earliest winged ancestors. Orders Odonata and Ephemeroptera form Paleoptera, which have outspread wings. The remaining orders have wings that can fold back over the abdomen (Neoptera). Superorder Orthopterodea includes orders Orthoptera, Isoptera, Plecoptera, Embioptera, and Dermaptera. Hemipterodea includes orders Zoraptera, Psocoptera, Hemiptera, Homoptera, Thysanoptera, Anoplura, and Mallophaga; and superorder Holometabola encompasses all holometabolous orders. Outgroups for this cladogram would be myriapods. |
The ancestral winged insect gave rise to three lines, which differed in their ability to flex their wings. Two of these (Odonata and Ephemeroptera) have outspread wings or hold their wings vertically over the abdomen. The other line has wings that can fold back horizontally over the abdomen. It branched into three groups, all of which were present by the Permian period. One group with hemimetabolous metamorphosis, chewing mouthparts, and cerci includes Orthoptera, Dermaptera, Isoptera, and Embioptera; another group with hemimetabolous metamorphosis and a tendency toward sucking mouthparts includes Thysanoptera, Hemiptera, and Homoptera and perhaps also Psocoptera, Zoraptera, Mallophaga, and Anoplura, although there is some disagreement among authorities about the last group. Insects with holometabolous metamorphosis have the most specialized life history, and these apparently form a clade.
The adaptive properties of insects have been stressed throughout this section. The directions and ranges of their adaptive radiation, both structurally and physiologically, have been amazingly varied. Whether it be in the area of habitat, feeding adaptations, means of locomotion, reproduction, or general mode of living, the adaptive achievements of insects are truly remarkable.
Support our developers




