Theories of Taxonomy
Theories of
Txonomy
A theory of taxonomy establishes the principles that we use to recognize and to rank taxonomic groups. There are two currently popular theories of taxonomy, (1) traditional evolutionary taxonomy and (2) phylogenetic systematics (cladistics). Both are based on evolutionary principles. We will see, however, that these two theories differ on how evolutionary principles are used. These differences have important implications for how we use a taxonomy to study the evolutionary process.
The relationship between a taxonomic group and a phylogenetic tree or cladogram is important for both of these theories. This relationship can take one of three forms: monophyly, paraphyly, or polyphyly (Figure 10-4). A taxon is monophyletic if it includes the most recent common ancestor of the group and all descendants of that ancestor (see Figure 10-4A). A taxon is paraphyletic if it includes the most recent common ancestor of all members of a group and some but not all of the descendants of that ancestor (see Figure 10-4B). A taxon is polyphyletic if it does not include the most recent common ancestor of all members of a group; this condition requires that the group has had at least two separate evolutionary origins, usually requiring independent evolutionary acquisition of similar features (see Figure 10-4C). Both evolutionary and cladistic taxonomy accept monophyletic groups and reject polyphyletic groups in their classifications. They differ on the acceptance of paraphyletic groups, however, and this difference has important evolutionary implications.
Traditional Evolutionary Taxonomy
Traditional evolutionary taxonomy incorporates two different evolutionary principles for recognizing and ranking higher taxa: (1) common descent and (2) amount of adaptive evolutionary change, as shown on a phylogenetic tree. Evolutionary taxa must have a single evolutionary origin, and must show unique adaptive features.
The mammalian paleontologist George Gaylord Simpson (Figure 10-5) was highly influential in developing and formalizing the procedures of evolutionary taxonomy. According to Simpson, a particular branch on the evolutionary tree is given the status of a higher taxon if it represents a distinct adaptive zone. Simpson describes an adaptive zone as “a characteristic reaction and mutual relationship between environment and organism, a way of life and not a place where life is led.” By entering a new adaptive zone through a fundamental change in organismal structure and behavior, an evolving population can use environmental resources in a completely new way.
A taxon that comprises a distinct adaptive zone is termed a grade. Simpson gives the example of penguins as a distinct adaptive zone within birds. The lineage immediately ancestral to all penguins underwent fundamental changes in the form of the body and wings to permit a switch from aerial to aquatic locomotion (Figure 10-6). Aquatic birds that can fly both in the air and underwater are somewhat intermediate in habitat, morphology, and behavior between aerial and aquatic adaptive zones. Nonetheless, the obvious modifications of the wings and body of penguins for swimming represent a new grade of organization. Penguins are therefore recognized as a distinct taxon within the birds, the family Spheniscidae. The broader the adaptive zone when fully occupied by a group of organisms, the higher the rank that the corresponding taxon is given.
Evolutionary taxa may be either monophyletic or paraphyletic. Recognition of paraphyletic taxa requires, however, that our taxonomies distort patterns of common descent. An evolutionary taxonomy of the anthropoid primates provides a good example (Figure 10-7). This taxonomy places humans (genus Homo) and their immediate fossil ancestors in the family Hominidae, and it places the chimpanzees (genus Pan), gorillas (genus Gorilla), and orangutans (genus Pongo) in the family Pongidae. However, the pongid genera Pan and Gorilla share more recent common ancestry with the Hominidae than they do with the remaining pongid genus, Pongo. This arrangement makes the family Pongidae paraphyletic because it does not include humans, who also descend from the most recent common ancestor of all pongids (see Figure 10- 7). Evolutionary taxonomists nonetheless recognize the pongid genera as a single, family-level grade of arboreal, herbivorous primates having limited mental capacity; in other words, they show the same family-level adaptive zone. Humans are terrestrial, omnivorous primates who have greatly expanded mental and cultural attributes, thereby comprising a distinct adaptive zone at the taxonomic level of the family. Unfortunately, if we want our taxa to constitute adaptive zones, we compromise our ability to present common descent in the most straightforward taxonomic manner.
Traditional evolutionary taxonomy has been challenged from two opposite directions. One challenge states that because phylogenetic trees can be very difficult to obtain, it is impractical to base our taxonomic system on common descent and adaptive evolution. We are told that our taxonomy should represent a more easily measured feature, the overall similarity of organisms evaluated without regard to phylogeny. This principle is known as phenetic taxonomy. Phenetic taxonomy did not have a strong impact on animal classification, and scientific interest in this approach is in decline. Despite the difficulties of reconstructing phylogeny, zoologists still consider this endeavor a central goal of their systematic work, and they are unwilling to compromise this goal for purposes of methodological simplicity.
Phylogenetic Systematics/Cladistics
A second and stronger challenge to evolutionary taxonomy is one known as phylogenetic systematics or cladistics. As the first name implies, this approach emphasizes the criterion of common descent and, as the second name implies, it is based on the cladogram of the group being classified. This approach to taxonomy was first proposed in 1950 by the German entomologist, Willi Hennig (Figure 10-8) and therefore is sometimes called “Hennigian systematics.” All taxa recognized by Hennig’s cladistic system must be monophyletic. We saw how evolutionary taxonomists’ recognition of the primate families Hominidae and Pongidae distorts genealogical relationships to emphasize adaptive uniqueness of the Hominidae. Because the most recent common ancestor of the paraphyletic family Pongidae is also an ancestor of the Hominidae, recognition of the Pongidae is incompatible with cladistic taxonomy. To avoid paraphyly, cladistic taxonomists have discontinued use of the traditional family Pongidae, placing chimpanzees, gorillas, and orangutans with humans in the family Hominidae. We adopt the cladistic classification in many sections.
The disagreement on the validity of paraphyletic groups may seem trivial at first, but its important consequences become clear when we discuss evolution. For example, claims that amphibians evolved from bony fish, that birds evolved from reptiles, or that humans evolved from apes may be made by an evolutionary taxonomist but are meaningless to a cladist. We imply by these statements that a descendant group (amphibians, birds, or humans) evolved from part of an ancestral group (bony fish, reptiles, and apes, respectively) to which the descendant does not belong. This usage automatically makes the ancestral group paraphyletic, and indeed bony fish, reptiles, and apes as traditionally recognized are paraphyletic groups. How are such paraphyletic groups recognized? Do they share distinguishing features that are not shared by the descendant group?
Paraphyletic groups are usually defined in a negative manner. They are distinguished only by features absent from a particular descendant group, because any traits that they share from their common ancestry are present also in the excluded descendants (unless secondarily lost). For example, apes are those “higher” primates that are not humans. Likewise, fish are those vertebrates that lack the distinguishing characteristics of tetrapods (amphibians and amniotes). What does it mean then to say that humans evolved from apes? To the evolutionary taxonomist, apes and humans are different adaptive zones or grades of organization; to say that humans evolved from apes states that bipedal, tailless organisms of large brain capacity evolved from arboreal, tailed organisms of smaller brain capacity. To the cladist, however, the statement that humans evolved from apes says essentially that humans evolved from something that they are not, a trivial statement that contains no useful information. To the cladist, any statement that a particular monophyletic group descends from a paraphyletic one is nothing more than a claim that the descendant group evolved from something that it is not. Extinct ancestral groups are always paraphyletic because they exclude a descendant that shares their most recent common ancestor. Although many such groups have been recognized by evolutionary taxonomists, none are recognized by cladists.
Zoologists often construct paraphyletic groups because they are interested in a terminal, monophyletic group (such as humans), and they want to ask questions about its ancestry. It is often convenient to lump together organisms whose features are considered approximately equally distant from the group of interest and to ignore their own unique features. It is significant in this regard that humans have never been placed in a paraphyletic group, whereas most other organisms have been. Apes, reptiles, fishes, and invertebrates are all terms that traditionally designate paraphyletic groups formed by combining various “side branches” that are found when human ancestry is traced backward through the tree of life. Such a taxonomy can give the erroneous impression that all of evolution is a progressive march toward humanity or, within other groups, a progressive march toward whatever species humans designate as being the most “advanced.” Such thinking is a relic of preDarwinian views that there is a linear scale of nature having “primitive” creatures at the bottom and humans near the top just below angels. Darwin’s theory of common descent states, however, that evolution is a branching process with no linear scale of increasing perfection along a single branch. Nearly every branch will contain its own combination of ancestral and derived features. In cladistics, this perspective is emphasized by recognizing taxa only by their own unique properties and not grouping organisms only because they lack the unique properties found in related groups.
Fortunately, there is a convenient way to express the common descent of groups without constructing paraphyletic taxa. It is done by finding what is called the sister group of the taxon of interest to us. Two different monophyletic taxa are termed sister groups if they share common ancestry with each other more recently than either one does with any other taxa. The sister group of humans appears to be chimpanzees, with gorillas forming a sister group to humans and chimpanzees combined. Orangutans are the sister group of a clade that includes humans, chimpanzees, and gorillas; gibbons form the sister group of the clade that includes orangutans, chimpanzees, gorillas, and humans (see Figure 10-7)
Current State of Animal Taxonomy
The formal taxonomy of animals that we use today was established using the principles of evolutionary systematics and has been revised recently in part using the principles of cladistics. Introduction of cladistic principles initially has the effect of replacing paraphyletic groups with monophyletic subgroups while leaving the remaining taxonomy mostly unchanged. A thorough revision of taxonomy along cladistic principles, however, will require profound changes, one of which almost certainly will be abandonment of the Linnaean ranks. In our coverage of animal taxonomy, we will try as much as possible to use taxa that are monophyletic and therefore consistent with criteria of both evolutionary and cladistic taxonomy. We will continue, however, to use Linnaean ranks. In some cases in which commonly recognized taxa are clearly paraphyletic grades, we will note this fact and suggest alternative taxonomic schemes that contain only monophyletic taxa.
In discussing patterns of descent, we will avoid statements such as “mammals evolved from reptiles” that imply paraphyly and will instead specify appropriate sister-group relationships. We will avoid referring to groups of organisms as being primitive, advanced, specialized, or generalized because all groups of animals contain combinations of primitive, advanced, specialized, and generalized features; these terms are best restricted to describing specific characteristics and not the group as a whole.
Revision of taxonomy according to cladistic principles can cause confusion. In addition to new taxonomic names, we see old ones used in unfamiliar ways. For example, cladistic use of “bony fishes” includes amphibians and amniotes (including reptilian groups, birds, and mammals) in addition to finned, aquatic animals that we normally term “fish.” Cladistic use of “reptiles” includes birds in addition to snakes, lizards, turtles, and crocodilians; however, it excludes some fossil forms, such as synapsids, that were traditionally placed in the Reptilia (see Reptilian Groups, Birds and Mammals). Taxonomists must be very careful to specify when using these seemingly familiar terms whether the traditional evolutionary taxa or newer cladistic taxa are being discussed.
A theory of taxonomy establishes the principles that we use to recognize and to rank taxonomic groups. There are two currently popular theories of taxonomy, (1) traditional evolutionary taxonomy and (2) phylogenetic systematics (cladistics). Both are based on evolutionary principles. We will see, however, that these two theories differ on how evolutionary principles are used. These differences have important implications for how we use a taxonomy to study the evolutionary process.
The relationship between a taxonomic group and a phylogenetic tree or cladogram is important for both of these theories. This relationship can take one of three forms: monophyly, paraphyly, or polyphyly (Figure 10-4). A taxon is monophyletic if it includes the most recent common ancestor of the group and all descendants of that ancestor (see Figure 10-4A). A taxon is paraphyletic if it includes the most recent common ancestor of all members of a group and some but not all of the descendants of that ancestor (see Figure 10-4B). A taxon is polyphyletic if it does not include the most recent common ancestor of all members of a group; this condition requires that the group has had at least two separate evolutionary origins, usually requiring independent evolutionary acquisition of similar features (see Figure 10-4C). Both evolutionary and cladistic taxonomy accept monophyletic groups and reject polyphyletic groups in their classifications. They differ on the acceptance of paraphyletic groups, however, and this difference has important evolutionary implications.
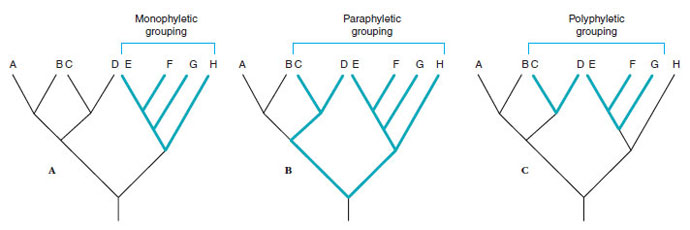 |
| Figure 10-4 Relationships between phylogeny and taxonomic groups illustrated for a hypothetical phylogeny of eight species (A through H). A, Monophyly—a monophyletic group contains the most recent common ancestor of all members of the group and all of its descendants. B, Paraphyly—a paraphyletic group contains the most recent common ancestor of all members of the group and some but not all of its descendants. C, Polyphyly—a polyphyletic group does not contain the most recent common ancestor of all members of the group, thereby requiring that the group have at least two separate phylogenetic origins. |
Traditional Evolutionary Taxonomy
 |
| Figure 10-5 George Gaylord Simpson (1902 to 1984) formulated the principles of evolutionary taxonomy. |
Traditional evolutionary taxonomy incorporates two different evolutionary principles for recognizing and ranking higher taxa: (1) common descent and (2) amount of adaptive evolutionary change, as shown on a phylogenetic tree. Evolutionary taxa must have a single evolutionary origin, and must show unique adaptive features.
The mammalian paleontologist George Gaylord Simpson (Figure 10-5) was highly influential in developing and formalizing the procedures of evolutionary taxonomy. According to Simpson, a particular branch on the evolutionary tree is given the status of a higher taxon if it represents a distinct adaptive zone. Simpson describes an adaptive zone as “a characteristic reaction and mutual relationship between environment and organism, a way of life and not a place where life is led.” By entering a new adaptive zone through a fundamental change in organismal structure and behavior, an evolving population can use environmental resources in a completely new way.
A taxon that comprises a distinct adaptive zone is termed a grade. Simpson gives the example of penguins as a distinct adaptive zone within birds. The lineage immediately ancestral to all penguins underwent fundamental changes in the form of the body and wings to permit a switch from aerial to aquatic locomotion (Figure 10-6). Aquatic birds that can fly both in the air and underwater are somewhat intermediate in habitat, morphology, and behavior between aerial and aquatic adaptive zones. Nonetheless, the obvious modifications of the wings and body of penguins for swimming represent a new grade of organization. Penguins are therefore recognized as a distinct taxon within the birds, the family Spheniscidae. The broader the adaptive zone when fully occupied by a group of organisms, the higher the rank that the corresponding taxon is given.
 |
| Figure 10-6 A, Penguin. B, Diving petrel. Penguins (avian family Spheniscidae) were recognized by George G. Simpson as a distinct adaptive zone within birds because of their adaptations for submarine flight. Simpson believed that the adaptive zone ancestral to penguins resembled that of diving petrels, which display adaptations for combined aerial and aquatic flight. Adaptive zones of penguins and diving petrels are distinct enough to be recognized taxonomically as different families within a common order (Ciconiiformes). |
Evolutionary taxa may be either monophyletic or paraphyletic. Recognition of paraphyletic taxa requires, however, that our taxonomies distort patterns of common descent. An evolutionary taxonomy of the anthropoid primates provides a good example (Figure 10-7). This taxonomy places humans (genus Homo) and their immediate fossil ancestors in the family Hominidae, and it places the chimpanzees (genus Pan), gorillas (genus Gorilla), and orangutans (genus Pongo) in the family Pongidae. However, the pongid genera Pan and Gorilla share more recent common ancestry with the Hominidae than they do with the remaining pongid genus, Pongo. This arrangement makes the family Pongidae paraphyletic because it does not include humans, who also descend from the most recent common ancestor of all pongids (see Figure 10- 7). Evolutionary taxonomists nonetheless recognize the pongid genera as a single, family-level grade of arboreal, herbivorous primates having limited mental capacity; in other words, they show the same family-level adaptive zone. Humans are terrestrial, omnivorous primates who have greatly expanded mental and cultural attributes, thereby comprising a distinct adaptive zone at the taxonomic level of the family. Unfortunately, if we want our taxa to constitute adaptive zones, we compromise our ability to present common descent in the most straightforward taxonomic manner.
Traditional evolutionary taxonomy has been challenged from two opposite directions. One challenge states that because phylogenetic trees can be very difficult to obtain, it is impractical to base our taxonomic system on common descent and adaptive evolution. We are told that our taxonomy should represent a more easily measured feature, the overall similarity of organisms evaluated without regard to phylogeny. This principle is known as phenetic taxonomy. Phenetic taxonomy did not have a strong impact on animal classification, and scientific interest in this approach is in decline. Despite the difficulties of reconstructing phylogeny, zoologists still consider this endeavor a central goal of their systematic work, and they are unwilling to compromise this goal for purposes of methodological simplicity.
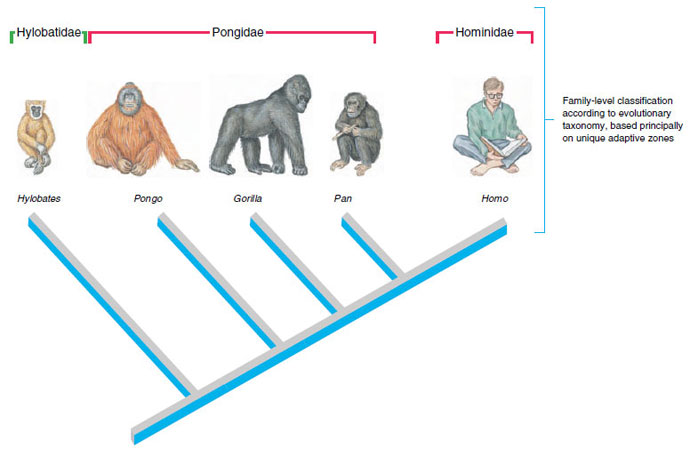 |
| Figure 10-7 Phylogeny and family-level classification of anthropoid primates. Evolutionary taxonomy groups the genera Gorilla, Pan, and Pongo into the paraphyletic family Pongidae because they share the same adaptive zone or grade of organization. Humans (genus Homo) are phylogenetically closer to Gorilla and Pan than any of these genera are to Pongo, but humans are placed in a separate family (Hominidae) because they represent a new grade of organization. Cladistic taxonomy discontinues recognition of the paraphyletic family Pongidae, consolidating Pongo, Gorilla, Pan, and Homo in the family Hominidae. |
 |
| Figure 10-8 Willi Hennig (1913 to 1976), German entomologist who formulated the principles of phylogenetic systematics/cladistics. |
Phylogenetic Systematics/Cladistics
A second and stronger challenge to evolutionary taxonomy is one known as phylogenetic systematics or cladistics. As the first name implies, this approach emphasizes the criterion of common descent and, as the second name implies, it is based on the cladogram of the group being classified. This approach to taxonomy was first proposed in 1950 by the German entomologist, Willi Hennig (Figure 10-8) and therefore is sometimes called “Hennigian systematics.” All taxa recognized by Hennig’s cladistic system must be monophyletic. We saw how evolutionary taxonomists’ recognition of the primate families Hominidae and Pongidae distorts genealogical relationships to emphasize adaptive uniqueness of the Hominidae. Because the most recent common ancestor of the paraphyletic family Pongidae is also an ancestor of the Hominidae, recognition of the Pongidae is incompatible with cladistic taxonomy. To avoid paraphyly, cladistic taxonomists have discontinued use of the traditional family Pongidae, placing chimpanzees, gorillas, and orangutans with humans in the family Hominidae. We adopt the cladistic classification in many sections.
The disagreement on the validity of paraphyletic groups may seem trivial at first, but its important consequences become clear when we discuss evolution. For example, claims that amphibians evolved from bony fish, that birds evolved from reptiles, or that humans evolved from apes may be made by an evolutionary taxonomist but are meaningless to a cladist. We imply by these statements that a descendant group (amphibians, birds, or humans) evolved from part of an ancestral group (bony fish, reptiles, and apes, respectively) to which the descendant does not belong. This usage automatically makes the ancestral group paraphyletic, and indeed bony fish, reptiles, and apes as traditionally recognized are paraphyletic groups. How are such paraphyletic groups recognized? Do they share distinguishing features that are not shared by the descendant group?
Paraphyletic groups are usually defined in a negative manner. They are distinguished only by features absent from a particular descendant group, because any traits that they share from their common ancestry are present also in the excluded descendants (unless secondarily lost). For example, apes are those “higher” primates that are not humans. Likewise, fish are those vertebrates that lack the distinguishing characteristics of tetrapods (amphibians and amniotes). What does it mean then to say that humans evolved from apes? To the evolutionary taxonomist, apes and humans are different adaptive zones or grades of organization; to say that humans evolved from apes states that bipedal, tailless organisms of large brain capacity evolved from arboreal, tailed organisms of smaller brain capacity. To the cladist, however, the statement that humans evolved from apes says essentially that humans evolved from something that they are not, a trivial statement that contains no useful information. To the cladist, any statement that a particular monophyletic group descends from a paraphyletic one is nothing more than a claim that the descendant group evolved from something that it is not. Extinct ancestral groups are always paraphyletic because they exclude a descendant that shares their most recent common ancestor. Although many such groups have been recognized by evolutionary taxonomists, none are recognized by cladists.
Zoologists often construct paraphyletic groups because they are interested in a terminal, monophyletic group (such as humans), and they want to ask questions about its ancestry. It is often convenient to lump together organisms whose features are considered approximately equally distant from the group of interest and to ignore their own unique features. It is significant in this regard that humans have never been placed in a paraphyletic group, whereas most other organisms have been. Apes, reptiles, fishes, and invertebrates are all terms that traditionally designate paraphyletic groups formed by combining various “side branches” that are found when human ancestry is traced backward through the tree of life. Such a taxonomy can give the erroneous impression that all of evolution is a progressive march toward humanity or, within other groups, a progressive march toward whatever species humans designate as being the most “advanced.” Such thinking is a relic of preDarwinian views that there is a linear scale of nature having “primitive” creatures at the bottom and humans near the top just below angels. Darwin’s theory of common descent states, however, that evolution is a branching process with no linear scale of increasing perfection along a single branch. Nearly every branch will contain its own combination of ancestral and derived features. In cladistics, this perspective is emphasized by recognizing taxa only by their own unique properties and not grouping organisms only because they lack the unique properties found in related groups.
Fortunately, there is a convenient way to express the common descent of groups without constructing paraphyletic taxa. It is done by finding what is called the sister group of the taxon of interest to us. Two different monophyletic taxa are termed sister groups if they share common ancestry with each other more recently than either one does with any other taxa. The sister group of humans appears to be chimpanzees, with gorillas forming a sister group to humans and chimpanzees combined. Orangutans are the sister group of a clade that includes humans, chimpanzees, and gorillas; gibbons form the sister group of the clade that includes orangutans, chimpanzees, gorillas, and humans (see Figure 10-7)
Current State of Animal Taxonomy
The formal taxonomy of animals that we use today was established using the principles of evolutionary systematics and has been revised recently in part using the principles of cladistics. Introduction of cladistic principles initially has the effect of replacing paraphyletic groups with monophyletic subgroups while leaving the remaining taxonomy mostly unchanged. A thorough revision of taxonomy along cladistic principles, however, will require profound changes, one of which almost certainly will be abandonment of the Linnaean ranks. In our coverage of animal taxonomy, we will try as much as possible to use taxa that are monophyletic and therefore consistent with criteria of both evolutionary and cladistic taxonomy. We will continue, however, to use Linnaean ranks. In some cases in which commonly recognized taxa are clearly paraphyletic grades, we will note this fact and suggest alternative taxonomic schemes that contain only monophyletic taxa.
In discussing patterns of descent, we will avoid statements such as “mammals evolved from reptiles” that imply paraphyly and will instead specify appropriate sister-group relationships. We will avoid referring to groups of organisms as being primitive, advanced, specialized, or generalized because all groups of animals contain combinations of primitive, advanced, specialized, and generalized features; these terms are best restricted to describing specific characteristics and not the group as a whole.
Revision of taxonomy according to cladistic principles can cause confusion. In addition to new taxonomic names, we see old ones used in unfamiliar ways. For example, cladistic use of “bony fishes” includes amphibians and amniotes (including reptilian groups, birds, and mammals) in addition to finned, aquatic animals that we normally term “fish.” Cladistic use of “reptiles” includes birds in addition to snakes, lizards, turtles, and crocodilians; however, it excludes some fossil forms, such as synapsids, that were traditionally placed in the Reptilia (see Reptilian Groups, Birds and Mammals). Taxonomists must be very careful to specify when using these seemingly familiar terms whether the traditional evolutionary taxa or newer cladistic taxa are being discussed.
Support our developers




