It is believed that one aspect of development and differentiation is the differential activation of various genes at different stages of development. Such a differential activation of developmental genes has actually been demonstrated in some cases. In recent years, mutations in specific genes have been identified which lead to failure of the development of specific organs. Such mutants have been studied in large number and have been very useful in identification, isolation and cloning of specific genes involved in developmental pathway. Some of the examples of differential activity of these genes and the mechanisms involved in its regulation were actually discussed in
Regulation of Gene Expression 1. Operon Circuits in Bacteria and other Prokaryotes,
Regulation of Gene Expression 2. Cricuit of Lytic Cycle and Lysogeny in Bacteriophages and
Regulation of Gene Expression 3. A Variety of Mechanisms in Eukaryotes. In this section, we will discuss cases, where differential gene activation has been demonstrated during development.
Gene regulation during early embryonic development
It has been demonstrated in several organisms (including
Drosophila, Xenopus and other amphibians, mouse, etc.), that just after fertilization, cell divisions are rapid and are controlled by maternally derived information in the cytoplasm. The zygotic nuclei do not exercise any genetic control in these early embryonic cell divisions, but start playing a role only after initial cell divisions are over. Techniques have now been developed which enable to locate the stage in development, at which the embryonic (or zyotic) nuclei start playing a role in the development. These approaches include the following :
Nuclear transplantation. Nuclear transplantation from sperms of one species into enucleated eggs of another species has been utilized to locate the stage at which paternal influence becomes obvious.
Location of stage for RNA synthesis. The stage at which some gene product (either RNA or protein) encoded by paternal genes becomes available, can be located by using drugs like actinomycin D, which would inhibit RNA synthesis on DNA template, causing lethality. The stage at which actinomycin D starts having lethal effect will suggest that the products of transcription or translation are essential for survival of the developing embryo. In amphibians, zygotic genes start functioning at the time of gastrulation. In mouse embryos, however, lethal effects of actinomycin D begin to appear around 9 or 16 cell stage of development.
Activation of specific genes in development. There are a number of genes, which act early in development and whose products are found in large quantities in differentiated cells thus facilitating their identification. In
Xenopus, until gastrulation (when differentiation of three similar cell layers occurs), maternal mRNA is found in good quantity and no new genes are activated. From gastrulae, mRNA could be isolated on poly-dT column, due to the presence of poly-A tails. Such mRNA contains both the maternal mRNA and zygotic mRNA and has been used for synthesis of cDNA through the use of reverse transcription.
This cDNA has been used for differential or subtractive hybridization with mRNA from unfertilized egg, so that cDNA not hybridized with this mRNA can be isolated as single stranded DNA and used for making double stranded cDNA. This double stranded cDNA has been used to make a library of upto 1,50,000 clones (many of them will be present repetitively in this library). This library should represent DNA sequences complementary to mRNA differentially synthesized at the gastrulation stage. When a number of these clones bound on filters were hybridized with radioactively labelled mRNA extracted from embryos at different stages of development, one could identify the number of genes and the degree of their expression at different developmental stages (consult next sections for techniques). Such experiments led to the following conclusions : (i) Three cDNA clones represent genes that are activated during gastrulation. (ii) Level of expression of different genes during gastrulation varies from gene to gene, (iii) Most of these individual mRNAs declined in abundance in later stages, although total mass of mRNA increases, suggesting that most of these genes are shut off by the tadpole stage.
Puffing patterns in giant chromosomes. The giant polytene chromosomes or salivary gland chromosomes of Diptera, is a system which has been utilized to demonstrate the visible differential gene activity. The swollen puffs and
Balbiani rings are the sites of RNA synthesis as demonstrated by incorporation of H
3 lebeled uracil at these sites, after incubation of larvae in this medium. The size and patterns of these puffs differ at different stages of development and indicate predictable differential gene activity. This has been demonstrated by giving heat shocks or treatment with steroid hormones, which lead to correlated changes in puffing patterns on the one hand and that of the appearance of specific mRNA and proteins on the other.
Use of markers in developmental genetics
Both phenotypic and genetic markers have been utilized in studies of development. For instance, by marking specific cells of an amphibian embryo, with visible carbon particles, movements of marked cells and their descendants during cell divisions could be traced during cell divisions. Using this technique,
fate maps could be prepared, showing the destinies of the descendants of particular cells in early stages of embryogenesis. Similarly, one may mark the cells genetically, by taking them from different sources, fuse them and then trace their fate during development.
Tetraparental mouse. Two mouse embryos (at 8 or 16 celled stage) each derived from two parents, can be fused together, after stripping off their surrounding membranes (using pronase enzyme). This may then be implanted in a host female, so that a tetraparental embryo will develop as a single mosaic individual. As an example,
Beatrice Mintz (a lady scientist) fused embryos (8-16 cells) from two strains, one having black fur and the other having white fur. The mosaic individual which developed, had bands of white and black fur circling the whole body. This suggested that the cells, whose descendants produced pigment, move after fusion of embryos and get lined up randomly in pairs, each member of a pair dividing to form a clonal sheet extending halfway around the body to meet the other sheet. Utilizing tetraparental mice, Mintz also resolved the controversy whether a multinucleate muscle cell originates due to a series of nuclear divisions without the division of cytoplasm or due to successive fusion of different myoblasts. Using genotypes with different forms,
ld-1a and
Id-1b of an enzyme (
isocitrate dehydrogenase)
, it could be shown that a striated muscle cell is formed by myoblast fusion, as evident from the formation of hybrid band (dimer) of the enzyme.
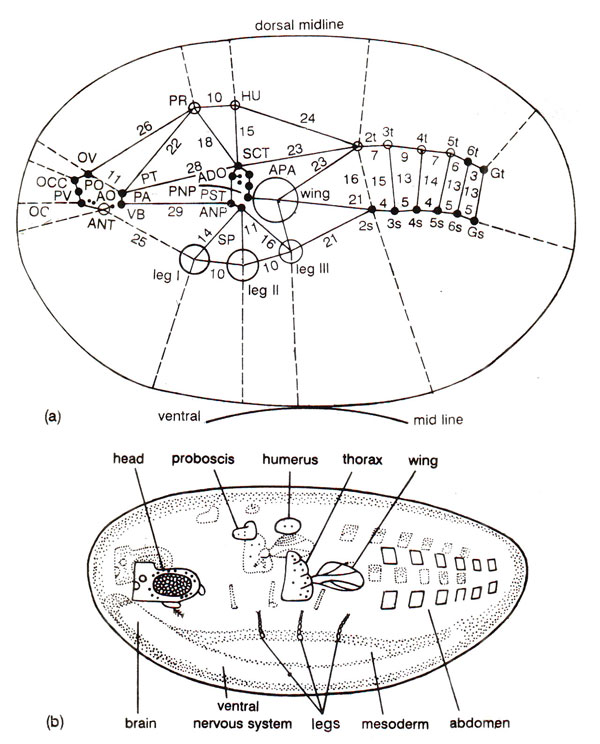
Fig. 38.2. (a) An embryonic fate map of adult structures in Drosophila ; the map shows foci for various structures and individual bristles; the distances are shown in map units called sturts (redrawn from Benzer, 1973). (b) A redrawn version of the above fate map, showing that the arrangement of structures in adult retains the pattern shown in the embryonic fate map (redrawn from Benzer, 1973).
Genetic mosaics in Drosophila. In
Drosophila also, genetic mosaics have been produced as a result of two different phenomena, namely
mitotic crossing over and
chromosome loss. Mitotic crossing over leads to the production of twin spots of homozygous tissues (see
Tetrad Analysis, Mitotic Recombination and Gene Conversion in Haploid Organisms (Fungi and Single Celled Algae)) and size of these spots depends on the stage at which mitotic crossing over occurred (earlier the crossing over, bigger is the size of spots). A study of a set of twin spots of different sizes would help to trace the cell lineage of an organ or a part of it. Atleast two cases are worth mentioning : (i) A heterozygote
wco/
w (
w = white eye) gave twin spots, which allowed tracing of cell lineage of lower half of the eye to eight early larval cells, (ii) A study of twin spots in a heterozygote,
yHw+/++ sn3 (
y = yellow body;
Hw = extra bristles along wing veins;
sn3 = short gnarled bristles) gave evidence that
Hw allele acts before the last 12 hours of larval life to imprint its phenotype irreversibly on the cell committed to form a wing.
A number of mutants in
Drosophila are now known, which have a tendency for chromosome loss giving rise to two types of tissues-one with full complement of chromosomes and the other with the deficiency of a specific chromosome. These tissues indicate that cells more closely related by cell division tend to stay together and therefore have been used for preparing
fate maps in
Drosophila (Fig. 38.2). Such fate maps have relationship with the actual spatial distribution of determined cells in the blastoderm.
Fig. 38.2. (a) An embryonic fate map of adult structures in Drosophila ; the map shows foci for various structures and individual bristles; the distances are shown in map units called sturts (redrawn from Benzer, 1973). (b) A redrawn version of the above fate map, showing that the arrangement of structures in adult retains the pattern shown in the embryonic fate map (redrawn from Benzer, 1973).




