Chromosome theory of inheritance suggested that Mendelian factors or genes were located on chromosomes. This theory was proved to hold good through a variety of experimental evidences. Since the chromosome complement in male and female gametes obtained from same individual would be similar, reciprocal crosses (♀A x ♂B : ♀B x ♂A) should give same results. The only exception to this expectation, earlier studied in
Sex Linked, Sex Influenced and Sex Limited Traits is sex linked inheritance. Differences in reciprocal crosses involving sex linked characters can be easily explained on the basis of transmission of sex chromosomes.
Since chromosomes divide in a very precise manner both during mitosis as well as during meiosis, it is easy to draw a parallelism between chromosomes and genes. Cytoplasm, however, does not divide in such a precise manner during cell division. Female gamete usually contributes more cytoplasm to the zygote.
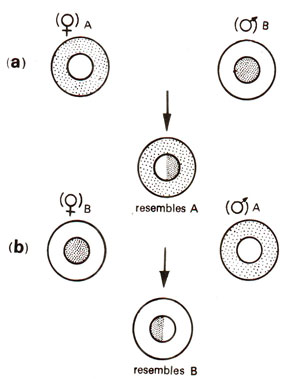
Fig. 18.1. Maternal inheritance showing difference in reciprocal crosses, (a) ♀A x ♂B, (b) ♀B x ♂A.
Consequently, for characters having cytoplasmic control, differences in reciprocal crosses would be observed. Inheritance in these cases would be mainly of maternal type as shown in Figure 18.1. As can be seen in the figure, if two strains A and B respectively having genotypes
AA and
BB and cytoplasms a and b are crossed reciprocally, we will get two hybrids
AB (a) and
AB (b) (cytoplasm is indicated in parentheses). In case of maternal effect,
AB (a) and
AB (b), despite having same nuclear genotype will differ.
AB (a) will resemble strain A or
AA (a) and
AB (b) will resemble strain B
or BB (b). Since such effects are solely produced by cytoplasm of the egg, they are described as maternal effects.
However, maternal effects are often produced due to effect of genes through cytoplasm. In other words, properties of cytoplasm depend on nuclear genes. Such cases can be distinguished from those, where extrachromosomal or cytoplasmic hereditary units are present and function either independently or in collaboration with nuclear genetic system. This is called
extrachromosomal or
cytoplasmic or
organellar inheritance, and is distinguished from maternal effects discussed above.
We know that in chromosomes, DNA is the sole genetic material and is the storehouse of genetic information. Discoveries of presence of DNA in cell organelles found outside the nucleus is a strong evidence to suggest that genetic information does exist in cytoplasm also. It will be seen in this section, that two important and essential organelles i.e. plastids (in plants only) and mitochondria located in cytoplasm carry DNA. These organelles control extrachromosomal inheritance in many cases through their DNA, which carries genetic information.
Fig. 18.1. Maternal inheritance showing difference in reciprocal crosses, (a) ♀A x ♂B, (b) ♀B x ♂A.
However, there are other cases, where cytoplasmic inheritance depends on extra-chromosomal particles, which are not essential for cell function and, therefore, may be present or absent. Such dispensable particles include
kappa in
Paramecium, and
sigma in
Drosophila. These particles are not only inherited but are also infective, since they can be introduced into new hosts without the need of actual process of reproduction. Further, the presence and reproduction of these infective particles may also depend on nuclear genes. Therefore, in this section we will discuss the following three kinds of examples: (i) Maternal effects depending indirectly on nuclear genes and involving no known cytoplasmic hereditary units, (ii) Cytoplasmic inheritance involving, dispensable and infective hereditary particles in cytoplasm, which may or may not depend on nuclear genes, (iii) Cytoplasmic inheritance involving essential organelles like chloroplasts and mitochondria




