Aluminum In Plant Tissues
Aluminum in Roots
Increasing aluminum levels in the medium tended to result in increasing aluminum concentrations in roots of aluminum accumulators or aluminum excluders (Table 16.3). Concentrations of aluminum in roots were 2- to 250-fold higher than those in shoots (Table 16.3). In red spruce, root aluminum concentrations associated with a 20% decrease in root biomass ranged from 1700 to 6000 mg Al kg-1 (78). Aluminum in roots is present mostly as precipitated hydroxy or phosphate compounds outside the root cells (86). As a result, it is difficult to use aluminum concentrations in roots as a measure of aluminum toxicity unless an effort is made to remove or prevent extracellularly precipitated and adsorbed aluminum. Alternatively, it might be possible to analyze aluminum concentrations in root apices alone as a measure of toxicity (189–191).Aluminum in Shoots
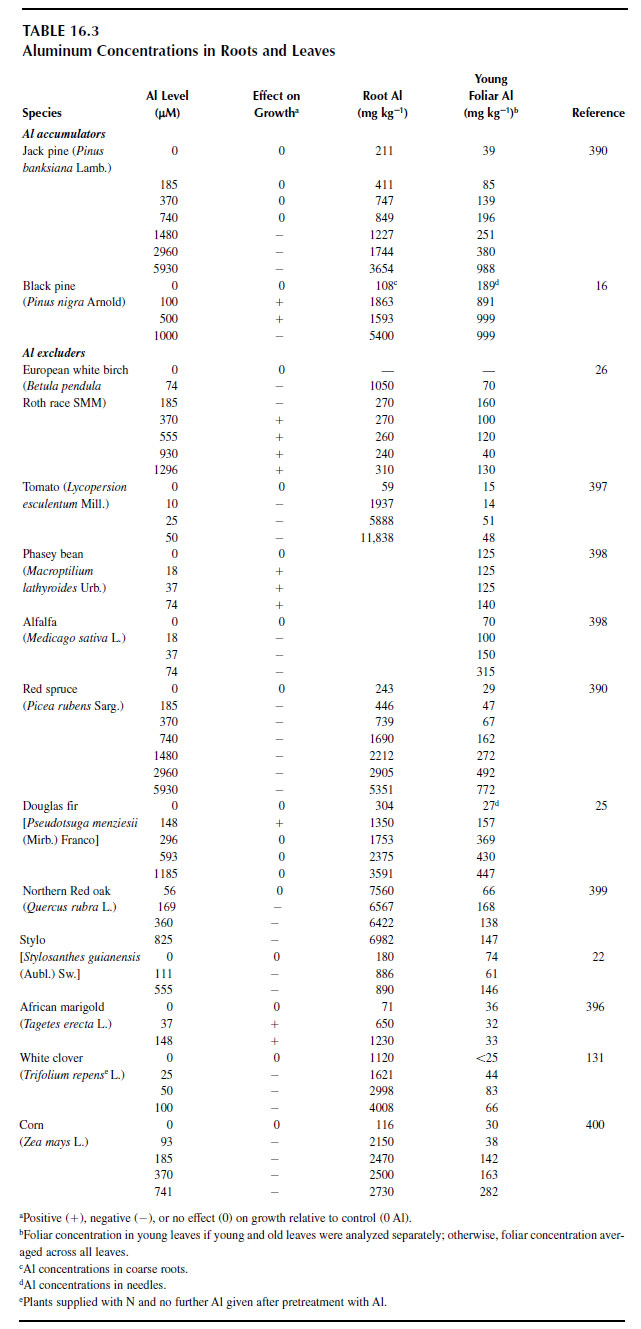
In accumulators, foliar aluminum concentrations of 65 tree species and 12 unidentified trees from an Indonesian rain forest ranged from 1 g kg-1 in delta tree (Aporusa spp. Blume, Euphorbiaceae) to 37 g kg-1 in Maschalocorymbosus corymbosus Bremek. (Rubiaceae) (13). Aluminum accumulators (Melastoma malabathricum L., Hydrangea macrophylla Ser., and Fagopyrum esculentum Moench.) exposed to increasing aluminum in solution showed increasing aluminum concentrations in leaves (22) (Figure 16.7). Facultative aluminum accumulators, jack pine (Pinus banksiana Lamb.) and loblolly pine (Pinus taeda L.), also had increasing foliar aluminum concentrations as solution aluminum increased (389) (Table 16.3).Efforts to establish critical aluminum concentrations for toxicity in plants generally have been unsuccessful (78,82,390). For example, foliar concentrations in red spruce associated with a 20% decrease in foliar biomass ranged from 70 to 250 mg kg-1 (78). Similarly, foliar aluminum concentrations in red oak associated with a 20% decrease in leaf biomass ranged from 93 to 188 mg kg-1 (391). Within slash pine families, aluminum sensitivity was correlated positively with foliar aluminum concentration; however, no such correlation was found within loblolly pine families (392).
In accumulators, internal complexation of aluminum by organic anions, silicate, or other ligands resulted in poor correlations between foliar aluminum concentrations and restrictions in biomass growth. Raynal et al. (78) reported the absence of any significant correlation between biomass response and foliar aluminum levels in Pinus species. In the case of aluminum excluders, aluminum concentrations in shoots do not increase with increasing aluminum levels in the medium until a toxic threshold is exceeded (328), again resulting in poor correlation between foliar aluminum levels and biomass response. For example, in rice and barley, only trace amounts of aluminum were found in leaves at solution aluminum levels up to 111 µM, then foliar aluminum concentrations increased as aluminum levels in solution increased to 555 µM (22) (Figure 16.7). Similarly, increasing solution aluminum levels from 0 to 620 µM had no effect on biomass growth of Western hemlock (Tsuga heterophylla Sarg.), then foliar aluminum concentrations decreased from 300 to 250 mg kg-1 when biomass was affected adversely by solution aluminum (393). In sugar maple (Acer saccharum Marsh.), aluminum concentrations in leaves increased from 50 to 200 mg kg-1 as aluminum levels in solution increased from 0 to 600 µM, but then foliar aluminum concentration dropped to 150 mg kg-1 when shoot growth was restricted at 1000 µM aluminum in solution (394). Other examples of a lack of correlation between aluminum-induced growth inhibition and foliar aluminum concentrations can be found in Table 16.3 (395–399).
 |
 |
| FIGURE 16.7 The pattern of increasing foliar aluminum concentrations with increasing solution aluminum differs in aluminum accumulator species (top) and aluminum excluder species (bottom) (From M. Osaki, T. Watanabe, T. Tadano. Beneficial effect of aluminum on growth of plants adapted to low pH soils. Soil Sci. Plant Nutr. 43:551–563, 1997.) |
Support our developers




