Glycerolipids and Fatty Acid Modification
Phosphatidic acid is generally metabolized by one of two enzymes. CDP-DAG
synthase, an enzyme found in ER, plastids and mitochondria, generates substrate for
production of phosphatidylglycerol, phosphatidylinositol, and phosphatidylserine. The other enzyme, phosphatidate phosphatase, releases DAG, a vital precursor of
PC, phosphatidylethanolamine and TAG, as well as sulfolipid and galactolipid. In
some plants,microsomal phosphatidate phosphatase suppliesDAG for both plastidial
and microsomal glycerolipid synthesis, while in others, separate plastidial and
microsomal isoforms contribute. Analysis of the phosphatase is complicated further
by isozymes involved in signaling and lipid catabolism. Based on work with developing
safflower seeds, Ichihara et al. proposed that an isoform used during oil
deposition moves between a cytosolic pool and the ER, depending on cytosolic
fatty acid concentrations (Ichihara
et al., 1990). This arrangement could allow feedforward
regulation of the TAG synthetic pathway initiated by the phosphatase.
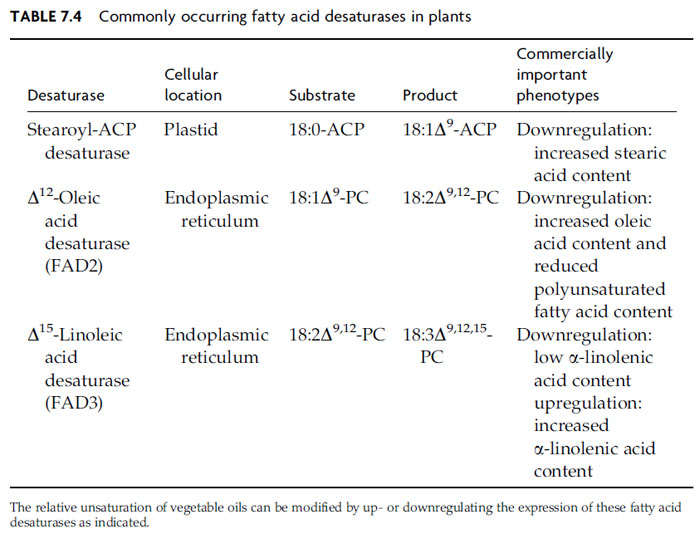
TAG composition can be radically affected by fatty acid modifications that take
place on glycerolipid substrates. As noted earlier, 18:1Δ
9 accounts for virtually all
of the unsaturated fatty acid exported by a typical plastid. Production of the
polyunsaturated fatty acids so common in vegetable oils involves a series of two
ER-localized desaturases that act on fatty acids esterified to either
sn-position of
PC or less prominent phospholipids (Fig. 7.4 and Table 7.4). The first enzyme, typically described as the Δ
12-oleic acid desaturase or
FAD2, inserts a double bond
12 carbons from the carboxyl end of esterified 18:1Δ
9, producing 18:2Δ
9,12 (linoleic
acid). This enzyme is sometimes referred to as the ω-6 desaturase, which indicates
that the double bond is inserted at the sixth carbon atom from the methyl end of
the 18:1Δ
9 substrate. A more careful analysis showed that this desaturase actually
references the site of double-bond insertion based on the position of the Δ
9 double
bond of its monounsaturated substrate (Schwartzbeck
et al., 2001). The second
enzyme, the D15-linoleic acid desaturase or FAD3, converts 18:2Δ
9,12 to 18:3Δ
9,12,15 (a-linolenic acid). As with
FAD2, this enzyme is sometimes referred to as the ω-3
desaturase, which indicates that the double bond is inserted at the third carbon
atom from the methyl end of its substrate. Engeseth and Stymne found that
FAD2 and FAD3 will also desaturate fatty acids that contain hydroxyl and epoxy groups
(Engeseth and Stymne, 1996). When determining insertion sites for new double
bonds, these enzymes appear to count the unusual functional groups as substitutes
for prior double bonds.
Again, the ER enzymes have plastidial counterparts, which act primarily on
glycolipid substrates.
FAD2 and FAD3 and the analogous plastidial desaturases
share eight conserved histidines arranged as H(X
3–4)H(X
7–41)H(X
2–3)HH(X
61–189)
H(X
2–3)HH, and it has been proposed that these histidines are associated with an
active site di-iron cluster (Shanklin and Cahoon, 1998). The same motif occurs in enzymes catalyzing a range of fatty acyl desaturation, hydroxylation, and
epoxidation reactions (Shanklin and Cahoon, 1998).