Altruistic Behavior and Kin Selection
Altruistic Behavior
and Kin Selection
If, as Darwin suggested, animals should behave selfishly and strive to produce as many offspring as possible, why do some animals help others at some risk to themselves? Why do some individuals show utmost cooperation with members of their social group and even forego breeding themselves? Why do some individuals appear to sacrifice themselves so that other members of their group can survive? Until the mid 1960s, scientists had trouble explaining in Darwinian terms how such altruistic behavior could persist in a population. Most instances of altruistic behaviors were explained using a groupselection argument. Group selectionists suggested that animals that helped others or that failed to mate did so for the benefit of the other members of the group. Therefore, such behaviors produce increased survivorship of groups whose members behaved altruistically. According to proponents of this argument, selection occurs at the level of the group, not at the level of the individual as Darwin suggested. However, the group-selection argument as originally proposed by V. C. Wynne- Edwards in 1962 has been rejected by the vast majority of behavioral ecologists for a number of reasons.
For example, if in a social group there were randomly distributed genes for a risky altruistic behavior, such as giving calls to warn others of predators, those lacking such genes would flourish. They would be warned with no risk to themselves; their chances of reproduction would be greater and, in time, the “selfish” alleles would eliminate the altruistic ones from the group’s gene pool.
In 1964, W. D. Hamilton, based largely on his studies of insects, proposed a new way to explain altruistic behavior by modifying Darwin’s original concept of fitness. He reasoned that fitness is measured not just by the number of offspring produced but by the increase or decrease in particular alleles in the gene pool. Thus, an individual may act altruistically, even at great risk, if it helps increase representation of its alleles in the gene pool. Alleles are shared by all relatives, including parents and offspring, brothers and sisters, cousins and other relations. Alleles that influence altruistic behavior among relatives would persist in future generations. Since the most closely related animals share the most genes by common descent, we expect that altruistic behavior would be most common among closely related individuals. Thus, if everything else were equal, brothers who on average share half their alleles would be more likely to aid one another than they would a cousin who shares on average only 25% of their alleles. Hamilton’s hypothesis based on this genetic explanation for altruism and cooperation is called kin selection. Essentially, kin selection is the selection of genes by individuals assisting the survival and reproduction of individuals who possess the same genes by common descent.
Hamilton’s hypothesis revolutionized evolutionary and behavioral biology. The main criterion of Darwinian fitness is the relative number of an individual’s alleles that are passed to future generations. Hamilton, however, developed the concept of inclusive fitness, which is the relative number of an individual’s alleles that are passed on to future generations either as a result of an individual’s own reproductive success or that of related individuals. Thus, kin selection and inclusive fitness may be able to explain many altruistic behaviors that have perplexed biologists for many years.
A good example of altruism and kin selection in nature is the remarkable cooperation and coordination among euscocial insects such as ants, bees, and wasps. Through haplodiploidy , where males are haploid and females are diploid, sisters are genetically related on average by 75% rather than 50% (Figure 38-20). Sisters are more closely related to each other than to their own daughters! Therefore, they cooperate with other members of their social group, forego breeding themselves and aid the queen to produce more sisters who are more closely related (75% related) than potential offspring (50% related).
Female Belding’s ground squirrels, found in the High Sierra of California, give alarm calls when a predator approaches (Figure 38-21). Alarm calling warns other members of the social group and is risky to the alarm caller. However, the benefits to alarm calling outweigh the risks because alarm callers are warning related individuals. Thus, alarm-calling behavior, even if it is risky, may be favored by selection if it increases inclusive fitness of the caller.
Kinship theory suggests that animals may evolve an ability to recognize categories of relatives so that cooperation or aid-giving behavior will be directed more efficiently toward relatives. Although kin recognition behavior was discussed by Hamilton, little was known about it until almost 20 years after he wrote his seminal papers. Through a number of experimental studies we now know that a variety of species can discriminate between kin and non-kin. Such species are found among invertebrates, including isopods and insects, and vertebrates, including fishes, frog and toad tadpoles, birds, squirrels, and monkeys. Some species can even discriminate between full siblings and half-siblings and between cousins and unrelated individuals. Thus, some species have a finely tuned ability to identify relatives of various degrees of relatedness. The cues used in kin recognition vary from species to species. Birds use vocalizations whereas many other species use chemical cues.
If, as Darwin suggested, animals should behave selfishly and strive to produce as many offspring as possible, why do some animals help others at some risk to themselves? Why do some individuals show utmost cooperation with members of their social group and even forego breeding themselves? Why do some individuals appear to sacrifice themselves so that other members of their group can survive? Until the mid 1960s, scientists had trouble explaining in Darwinian terms how such altruistic behavior could persist in a population. Most instances of altruistic behaviors were explained using a groupselection argument. Group selectionists suggested that animals that helped others or that failed to mate did so for the benefit of the other members of the group. Therefore, such behaviors produce increased survivorship of groups whose members behaved altruistically. According to proponents of this argument, selection occurs at the level of the group, not at the level of the individual as Darwin suggested. However, the group-selection argument as originally proposed by V. C. Wynne- Edwards in 1962 has been rejected by the vast majority of behavioral ecologists for a number of reasons.
For example, if in a social group there were randomly distributed genes for a risky altruistic behavior, such as giving calls to warn others of predators, those lacking such genes would flourish. They would be warned with no risk to themselves; their chances of reproduction would be greater and, in time, the “selfish” alleles would eliminate the altruistic ones from the group’s gene pool.
In 1964, W. D. Hamilton, based largely on his studies of insects, proposed a new way to explain altruistic behavior by modifying Darwin’s original concept of fitness. He reasoned that fitness is measured not just by the number of offspring produced but by the increase or decrease in particular alleles in the gene pool. Thus, an individual may act altruistically, even at great risk, if it helps increase representation of its alleles in the gene pool. Alleles are shared by all relatives, including parents and offspring, brothers and sisters, cousins and other relations. Alleles that influence altruistic behavior among relatives would persist in future generations. Since the most closely related animals share the most genes by common descent, we expect that altruistic behavior would be most common among closely related individuals. Thus, if everything else were equal, brothers who on average share half their alleles would be more likely to aid one another than they would a cousin who shares on average only 25% of their alleles. Hamilton’s hypothesis based on this genetic explanation for altruism and cooperation is called kin selection. Essentially, kin selection is the selection of genes by individuals assisting the survival and reproduction of individuals who possess the same genes by common descent.
Hamilton’s hypothesis revolutionized evolutionary and behavioral biology. The main criterion of Darwinian fitness is the relative number of an individual’s alleles that are passed to future generations. Hamilton, however, developed the concept of inclusive fitness, which is the relative number of an individual’s alleles that are passed on to future generations either as a result of an individual’s own reproductive success or that of related individuals. Thus, kin selection and inclusive fitness may be able to explain many altruistic behaviors that have perplexed biologists for many years.
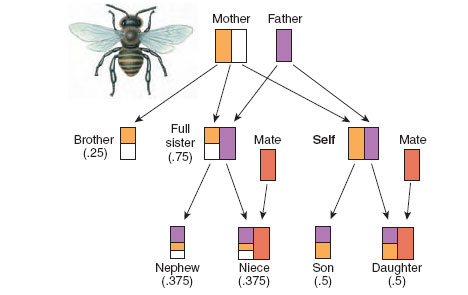 |
| Figure 38-20 Haplodiploidy in honey bees, showing degrees of relatedness of a female worker bee (labeled SELF) to individuals she might raise. In honey bees, as in other haplodiploid animals, diploid females develop from fertilized eggs,and males develop from unfertilized eggs. Each daughter of a male gets all his genes (purple bar) and full sisters receive an identical one half of their genome from the same father. Open bars represent other, unrelated alleles. Because full sisters also share half the genesthey receive from their common mother (yellow bar), the relatedness of SELF to a full sister is 0.75,the average of 0.5 and 1.0. (In a diploid-diploid system as in humans,the relatedness of siblings is 0.5 because both paternally and maternallyinherited genes have a 50% chance of being present in a sibling.)Note that relatedness of female workers to a brother is only 0.25, because brothers are fatherless. |
A good example of altruism and kin selection in nature is the remarkable cooperation and coordination among euscocial insects such as ants, bees, and wasps. Through haplodiploidy , where males are haploid and females are diploid, sisters are genetically related on average by 75% rather than 50% (Figure 38-20). Sisters are more closely related to each other than to their own daughters! Therefore, they cooperate with other members of their social group, forego breeding themselves and aid the queen to produce more sisters who are more closely related (75% related) than potential offspring (50% related).
 |
| Figure 38-21 Belding’s ground squirrel, Spermophilus beldingi,gives an alarm call to warn of the approach of a predator. This risky behaviorendangers the callers more than noncallers |
Female Belding’s ground squirrels, found in the High Sierra of California, give alarm calls when a predator approaches (Figure 38-21). Alarm calling warns other members of the social group and is risky to the alarm caller. However, the benefits to alarm calling outweigh the risks because alarm callers are warning related individuals. Thus, alarm-calling behavior, even if it is risky, may be favored by selection if it increases inclusive fitness of the caller.
Kinship theory suggests that animals may evolve an ability to recognize categories of relatives so that cooperation or aid-giving behavior will be directed more efficiently toward relatives. Although kin recognition behavior was discussed by Hamilton, little was known about it until almost 20 years after he wrote his seminal papers. Through a number of experimental studies we now know that a variety of species can discriminate between kin and non-kin. Such species are found among invertebrates, including isopods and insects, and vertebrates, including fishes, frog and toad tadpoles, birds, squirrels, and monkeys. Some species can even discriminate between full siblings and half-siblings and between cousins and unrelated individuals. Thus, some species have a finely tuned ability to identify relatives of various degrees of relatedness. The cues used in kin recognition vary from species to species. Birds use vocalizations whereas many other species use chemical cues.
Support our developers




