Animal Body Plans
Animal Body Plans
As mentioned in the prologue in the above topics, the diversity of animal body form is constrained by evolutionary history, habitat, and way of life. Although a worm that adopts a parasitic life in the intestine of a vertebrate looks and functions very differently from a free-living member of the same group, both share distinguishing hallmarks of their phylum. We consider here the limited number of basic body plans that underlie diversity of animal form and we examine common architectural themes that animals share.
Major evolutionary innovations in the forms of animals include multicellularity, bilateral symmetry, “tubewithin- a-tube” plan, and eucoelomate (true coelom) body plan. The evolutionary distributions of these body plans are shown in Figure 9-9.
Animal Symmetry
Symmetry refers to balanced proportions, or correspondence in size and shape of parts on opposite sides of a median plane.
Spherical symmetry means that any plane passing through the center divides the body into equivalent, or mirrored, halves (Figure 9-10, top left). This type of symmetry is found chiefly among some unicellular forms and is rare in animals. Spherical forms are best suited for floating and rolling.
Radial symmetry (Figure 9-10, top right) applies to forms that can be divided into similar halves by more than two planes passing through the longitudinal axis. These are tubular, vase, or bowl shapes found in some sponges and in hydras, jellyfish, sea urchins, and related groups, in which one end of the longitudinal axis is usually the mouth. A variant form is biradial symmetry in which, because of some part that is single or paired rather than radial, only two planes passing through the longitudinal axis produce mirrored halves. Sea walnuts (phylum Ctenophora), which are more or less globular in form but have a pair of tentacles, are an example. Radial and biradial animals are usually sessile, freely floating, or weakly swimming. Radial animals, with no front or back end, can interact with their environment in all directions—an advantage to sessile forms with feeding structures arranged to snare prey approaching from any direction.
The two phyla that are primarily radial, Cnidaria and Ctenophora, are called the Radiata. Echinoderms (sea stars and their kin) are primarily bilateral animals (their larvae are bilateral) that have become secondarily radial as adults.
Bilateral symmetry applies to
animals that can be divided along a
sagittal plane into two mirrored portions—
right and left halves (Figure
9-10, bottom). The appearance of
bilateral symmetry in animal evolution
was a major advancement, because
bilateral animals are much better fitted
for directional (forward) movement
than are radially symmetrical animals.
Bilateral animals form a monophyletic
group of phyla called the Bilateria.
Bilateral symmetry is strongly associated
with cephalization, discussed
below.
Some convenient terms used for locating regions of bilaterally-symmetrical animals (Figure 9-11) are anterior, used to designate the head end; posterior, the opposite or tail end; dorsal, the back side; and ventral, the front or belly side. Medial refers to the midline of the body; lateral, to the sides. Distal parts are far from the middle of the body; proximal parts are nearer. A frontal plane (sometimes called coronal plane) divides a bilateral body into dorsal and ventral halves by running through the anteroposterior axis and the right-left axis at right angles to the sagittal plane, the plane dividing an animal into right and left halves. A transverse plane (also called a cross section) would cut through a dorsoventral and a right-left axis at right angles to both the sagittal and frontal planes and would result in anterior and posterior portions (Figure 9-11).
In vertebrates pectoral
refers to the chest region or the area
supported by the forelegs, and pelvic
refers to the hip region or the area
supported by the hind legs.
Body Cavities
Bilateral animals can be grouped according to the presence and type of body cavity (Figure 9-12). A major evolutionary innovation appearing within the Bilateria is the coelom, a fluid-filled space that surrounds the gut. The coelom provides a tubewithin- a-tube arrangement (Figure 9-12) that allows much greater flexibility of the body cavity. The coelom also provides space for visceral organs and permits greater size and complexity by exposing more cells to surface exchange. The fluidfilled coelom additionally serves as a hydrostatic skeleton in some forms, especially many worms, aiding in such activities as movement and burrowing.
As shown in Figure 9-9, the presence or absence of a coelom is a key determinant in the evolutionary advancement of the Bilateria.
Acoelomate Bilateria
Many bilateral animals do not have a true coelom. In fact, flatworms and a few others have no body cavity surrounding the gut (Figure 9-12, top). The region between the ectodermal epidermis and the endodermal digestive tract is completely filled with mesoderm in the form of a spongy mass of space-filling cells called parenchyma. Parenchyma is derived from an inwandering of ectodermal cells from the general surface of the early embryo. In at least some acoelomates, the parenchymal cells are cell bodies of muscle cells.
Pseudocoelomate Bilateria
Nematodes and several other phyla have a cavity surrounding the gut, but it is not lined with mesodermal peritoneum. The cavity is derived from the blastocoel of the embryo and represents a persistent blastocoel. This type of body cavity is called a pseudocoel, and its possessors also have a tubewithin- a-tube arrangement (Figure 9-12, center).
Eucoelomate Bilateria
The remaining bilateral animals possess
a true coelom lined with mesodermal
peritoneum (Figure 9-12, bottom). The true coelom arises within the mesoderm
itself and may be formed by one of two
methods, schizocoelous or enterocoelous
development (Figure 9-13), or
by modifying these methods. The two
terms are descriptive, for schizo comes
from the Greek schizein, to split; entero is derived from the Greek enteron,
meaning gut; and coelous comes from
the Greek koilos, meaning hollow or
cavity. In schizocoelous formation the
coelom arises, as the word implies, from
splitting of mesodermal bands that originate
from cells in the blastopore region.
(Mesoderm is one of three primary germ
layers that appear very early in the
development of all bilateral animals,
lying between the innermost endoderm
and outermost ectoderm, and Figure
8-24). In enterocoelous formation
the coelom comes from pouches
of the archenteron, or primitive gut.
Once development is complete, the results of schizocoelous and enterocoelous formations are indistinguishable. Both give rise to a true coelom lined with mesodermal peritoneum (Gr. peritonaios, stretched around) and having mesenteries in which visceral organs are suspended.
Metamerism (Segmentation)
Metamerism is a serial repetition of similar body segments along the longitudinal axis of the body. Each segment is called a metamere, or somite. In forms such as earthworms and other annelids, in which metamerism is most clearly represented, the segmental arrangement includes both external and internal structures of several systems. There is repetition of muscles, blood vessels, nerves, and setae of locomotion. Some other organs, such as those of sex, may be repeated in only a few somites. Evolutionary changes have obscured much of the segmentation in many animals, including humans.
True metamerism is found in only three phyla: Annelida, Arthropoda, and Chordata (Figure 9-14), although superficial segmentation of ectoderm and body wall may be found among many diverse groups of animals.
Cephalization
Differentiation of a head is called cephalization and is found chiefly in bilaterally symmetrical animals. The concentration of nervous tissue and sense organs in the head bestows obvious advantages to an animal moving through its environment head first. This is the most efficient positioning of organs for sensing the environment and responding to it. Usually the mouth of the animal is located on the head as well, since so much of an animal’s activity is concerned with procuring food. Cephalization is always accompanied by differentiation along an anteroposterior axis (polarity). Polarity usually involves gradients of activities between limits, such as between anterior and posterior ends.
As mentioned in the prologue in the above topics, the diversity of animal body form is constrained by evolutionary history, habitat, and way of life. Although a worm that adopts a parasitic life in the intestine of a vertebrate looks and functions very differently from a free-living member of the same group, both share distinguishing hallmarks of their phylum. We consider here the limited number of basic body plans that underlie diversity of animal form and we examine common architectural themes that animals share.
Major evolutionary innovations in the forms of animals include multicellularity, bilateral symmetry, “tubewithin- a-tube” plan, and eucoelomate (true coelom) body plan. The evolutionary distributions of these body plans are shown in Figure 9-9.
Animal Symmetry
Symmetry refers to balanced proportions, or correspondence in size and shape of parts on opposite sides of a median plane.
Spherical symmetry means that any plane passing through the center divides the body into equivalent, or mirrored, halves (Figure 9-10, top left). This type of symmetry is found chiefly among some unicellular forms and is rare in animals. Spherical forms are best suited for floating and rolling.
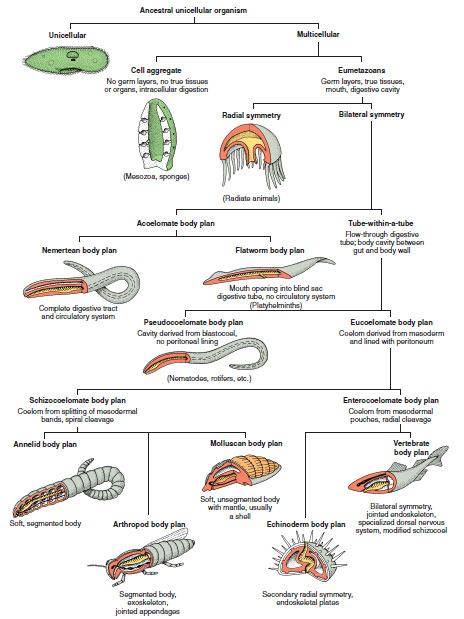 |
| Figure 9-9 Architectural patterns of animals. These basic body plans have been variously modified during evolutionary descent to fit animals to a great variety of habitats. Ectoderm is shown in gray, mesoderm in red, and endoderm in yellow. |
Radial symmetry (Figure 9-10, top right) applies to forms that can be divided into similar halves by more than two planes passing through the longitudinal axis. These are tubular, vase, or bowl shapes found in some sponges and in hydras, jellyfish, sea urchins, and related groups, in which one end of the longitudinal axis is usually the mouth. A variant form is biradial symmetry in which, because of some part that is single or paired rather than radial, only two planes passing through the longitudinal axis produce mirrored halves. Sea walnuts (phylum Ctenophora), which are more or less globular in form but have a pair of tentacles, are an example. Radial and biradial animals are usually sessile, freely floating, or weakly swimming. Radial animals, with no front or back end, can interact with their environment in all directions—an advantage to sessile forms with feeding structures arranged to snare prey approaching from any direction.
The two phyla that are primarily radial, Cnidaria and Ctenophora, are called the Radiata. Echinoderms (sea stars and their kin) are primarily bilateral animals (their larvae are bilateral) that have become secondarily radial as adults.
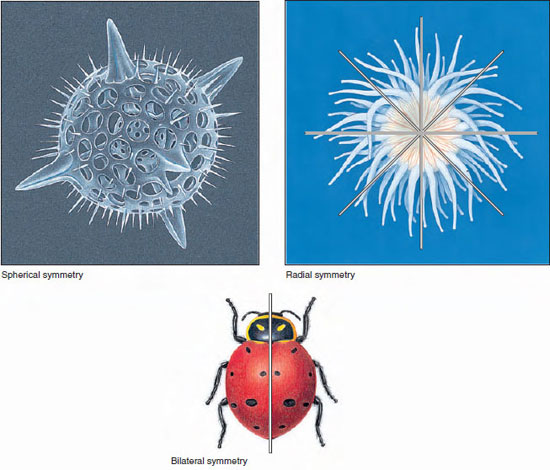 |
| Figure 9-10 Animal symmetry. Illustrated are animals showing spherical, radial, and bilateral symmetry. |
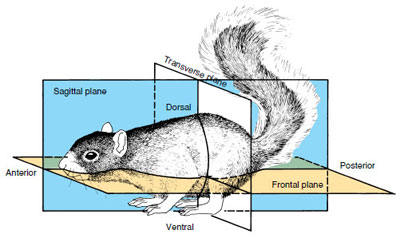 |
| Figure 9-11 The planes of symmetry as illustrated by a bilaterally symmetrical animal. |
Some convenient terms used for locating regions of bilaterally-symmetrical animals (Figure 9-11) are anterior, used to designate the head end; posterior, the opposite or tail end; dorsal, the back side; and ventral, the front or belly side. Medial refers to the midline of the body; lateral, to the sides. Distal parts are far from the middle of the body; proximal parts are nearer. A frontal plane (sometimes called coronal plane) divides a bilateral body into dorsal and ventral halves by running through the anteroposterior axis and the right-left axis at right angles to the sagittal plane, the plane dividing an animal into right and left halves. A transverse plane (also called a cross section) would cut through a dorsoventral and a right-left axis at right angles to both the sagittal and frontal planes and would result in anterior and posterior portions (Figure 9-11).
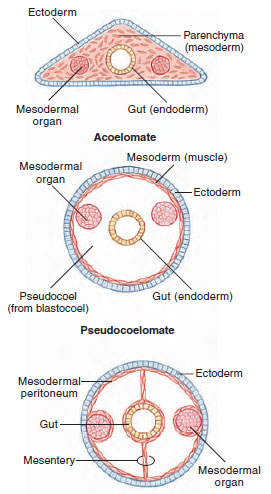 |
| Figure 9-12 Acoelomate, pseudocoelomate, and eucoelomate body plans. |
Body Cavities
Bilateral animals can be grouped according to the presence and type of body cavity (Figure 9-12). A major evolutionary innovation appearing within the Bilateria is the coelom, a fluid-filled space that surrounds the gut. The coelom provides a tubewithin- a-tube arrangement (Figure 9-12) that allows much greater flexibility of the body cavity. The coelom also provides space for visceral organs and permits greater size and complexity by exposing more cells to surface exchange. The fluidfilled coelom additionally serves as a hydrostatic skeleton in some forms, especially many worms, aiding in such activities as movement and burrowing.
As shown in Figure 9-9, the presence or absence of a coelom is a key determinant in the evolutionary advancement of the Bilateria.
Acoelomate Bilateria
Many bilateral animals do not have a true coelom. In fact, flatworms and a few others have no body cavity surrounding the gut (Figure 9-12, top). The region between the ectodermal epidermis and the endodermal digestive tract is completely filled with mesoderm in the form of a spongy mass of space-filling cells called parenchyma. Parenchyma is derived from an inwandering of ectodermal cells from the general surface of the early embryo. In at least some acoelomates, the parenchymal cells are cell bodies of muscle cells.
Pseudocoelomate Bilateria
Nematodes and several other phyla have a cavity surrounding the gut, but it is not lined with mesodermal peritoneum. The cavity is derived from the blastocoel of the embryo and represents a persistent blastocoel. This type of body cavity is called a pseudocoel, and its possessors also have a tubewithin- a-tube arrangement (Figure 9-12, center).
Eucoelomate Bilateria
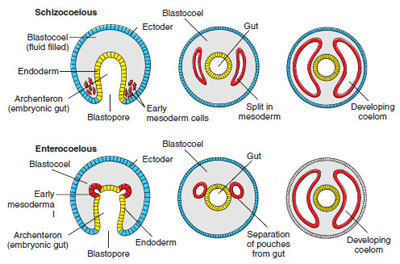 |
| Figure 9-13 Types of mesoderm and coelom formation. In schizocoelous formation, the mesoderm originates from the wall of the archenteron near the blastopore and proliferates into a band of tissue that splits to form the coelom. In enterocoelous formation, most mesoderm originates as a series of pouches from the archenteron; these pinch off and enlarge to form the coelom. In both formations, the coeloms expand to obliterate the blastocoel. |
Once development is complete, the results of schizocoelous and enterocoelous formations are indistinguishable. Both give rise to a true coelom lined with mesodermal peritoneum (Gr. peritonaios, stretched around) and having mesenteries in which visceral organs are suspended.
Metamerism (Segmentation)
Metamerism is a serial repetition of similar body segments along the longitudinal axis of the body. Each segment is called a metamere, or somite. In forms such as earthworms and other annelids, in which metamerism is most clearly represented, the segmental arrangement includes both external and internal structures of several systems. There is repetition of muscles, blood vessels, nerves, and setae of locomotion. Some other organs, such as those of sex, may be repeated in only a few somites. Evolutionary changes have obscured much of the segmentation in many animals, including humans.
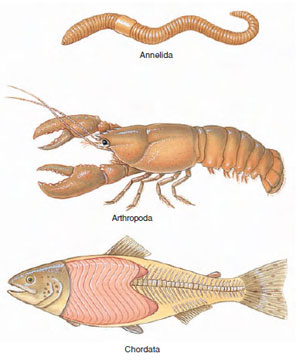 |
| Figure 9-14
Segmented phyla. These three phyla have all made use of an important principle in nature: metamerism, or repetition of structural units. Segmentation in annelids and arthropods is homologous, but chordates may have derived their segmentation independently. Segmentation brings more varied specialization because segments, especially in arthropods, have become modified for different functions. |
True metamerism is found in only three phyla: Annelida, Arthropoda, and Chordata (Figure 9-14), although superficial segmentation of ectoderm and body wall may be found among many diverse groups of animals.
Cephalization
Differentiation of a head is called cephalization and is found chiefly in bilaterally symmetrical animals. The concentration of nervous tissue and sense organs in the head bestows obvious advantages to an animal moving through its environment head first. This is the most efficient positioning of organs for sensing the environment and responding to it. Usually the mouth of the animal is located on the head as well, since so much of an animal’s activity is concerned with procuring food. Cephalization is always accompanied by differentiation along an anteroposterior axis (polarity). Polarity usually involves gradients of activities between limits, such as between anterior and posterior ends.
Support our developers




