The Hierarchical Organization of Animal Complexity
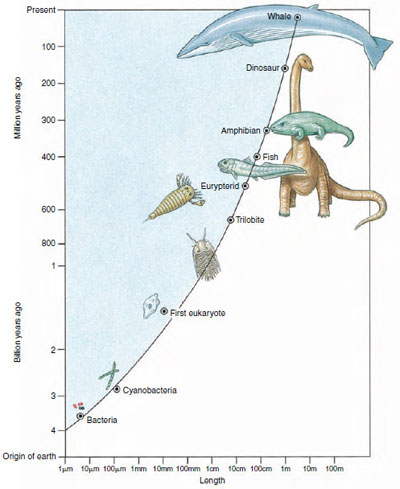 |
| Figure 9-1 Graph showing the evolution of size (length) increase in organisms at different periods of life on earth. Note that both scales are logarithmic. |
Among the different unicellular and metazoan groups, we can recognize five major grades of organization (Table 9-1). Each grade is more complex than the one before, and builds on it in a hierarchical manner.
The unicellular protozoa groups are the simplest animal-like organisms. They are nonetheless complete organisms that perform all of the basic functions of life as seen in the more complex animals. Within the confines of their cell, they show remarkable organization and division of labor, possessing distinct supportive structures, locomotor devices, fibrils, and simple sensory structures. The diversity observed among unicellular organisms is achieved by varying the architectural patterns of subcellular structures, organelles, and the cell as a whole (Protozoan Groups).
The metazoa, or multicellular animals, evolved greater structural complexity by combining cells into larger units. A metazoan cell is a specialized part of the whole organism and, unlike a protozoan cell, it is not capable of independent existence. Cells of a multicellular organism are specialized for performing the various tasks accomplished by subcellular elements in unicellular forms. The simpleset metazoans show the cellular grade of organization in which cells demonstrate division of labor but are not strongly associated to perform a specific collective function (Table 9-1). In the more complex tissue grade, similar cells are grouped together and perform their common functions as a highly coordinated unit. In animals of the tissueorgan grade of organization, tissues are assembled into still larger functional units called organs. Usually one type of tissue carries the burden of an organ’s chief function, as muscle tissue does in the heart; other tissues— epithelial, connective, and nervous— perform supportive roles.
Most metazoa (nemerteans and all more structurally complex phyla) have an additional level of complexity in which different organs operate together as organ systems. Eleven different kinds of organ systems are observed in metazoans: skeletal, muscular, integumentary, digestive, respiratory, circulatory, excretory, nervous, endocrine, immune, and reproductive. The great evolutionary diversity of these organ systems is covered in Acoelomate Animals, Pseudocoelomate Animals , Molluscs , Segmented Worms, Arthropods , Aquatic Mandibulates, Terrestrial Mandibulates, Lesser Protostomes , Lophophorate Animals , Echinoderms , Chaetognaths and Hemichordates, Chordates , Fishes, Early Tetrapods and Modern Amphibians, Reptilian Groups, Birds and Mammals .
 |
Complexity and Body Size
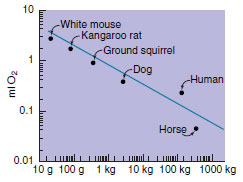 |
| Figure 9-2 Net cost of running for mammals of various sizes. Each point represents the cost (measured in rate of oxygen consumption) of moving 1 g of body over 1 km. The cost decreases with increasing body size. |
The opening essay suggests that size is a major consideration in the design of animals. The most complex grades of metazoan organization permit and to some extent even promote the evolution of large body size (Figure 9-1). Large size confers several important physical and ecological consequences for the organism. As animals become larger, the body surface increases much more slowly than body volume because surface area increases as the square of body length (length2), whereas volume (and therefore mass) increases as the cube of body length (length3). In other words, a large animal will have less surface area relative to its volume than will a small animal of the same shape. The surface area of a large animal may be inadequate for respiration and nutrition by cells located deep within the body. There are two possible solutions to this problem. One solution is to fold or invaginate the body surface to increase the surface area or, as exploited by the flatworms, flatten the body into a ribbon or disc so that no internal space is far from the surface. This solution allows the body to become large without internal complexity. However, most large animals adopted a second solution; they developed internal transport systems to shuttle nutrients, gases, and waste products between the cells and the external environment.
Larger size buffers the animal against environmental fluctuations; it provides greater protection against predation and enhances offensive tactics; and it permits a more efficient use of metabolic energy. A large mammal uses more oxygen than a small mammal, but the cost of maintaining its body temperature is less per gram of weight for the large mammal than for a small one. A large mammal uses more oxygen in running than a small mammal, but the energy cost of moving 1 g of its body over a given distance is much less for a large mammal than for a small one (Figure 9-2). For all of these reasons, ecological opportunities of larger animals are very different from those of small ones. In subsequent sections we will describe the extensive adaptive radiations observed in taxa of large animals.
Support our developers




