Class Chondrichthyes: Cartilaginous Fishes
Class Chondrichthyes:
Cartilaginous Fishes
There are nearly 850 living species in the class Chondrichthyes, an ancient, compact, and highly developed group. Although a much smaller and less diverse assemblage than the bony fishes, their impressive combination of well-developed sense organs, powerful jaws and swimming musculature, and predaceous habits ensures them a secure and lasting place in the aquatic community. One of their distinctive features is their cartilaginous skeleton. Although calcification may be extensive in their skeletons, bone is entirely absent throughout the class—a curious evolutionary feature, since the Chondrichthyes are derived from ancestors having well-developed bone. Almost all chondrichthyans are marine; only 28 species live primarily in fresh water.
With the exception of whales, sharks include the largest living vertebrates. The larger sharks may reach 12 m in length. Dogfish sharks so widely studied in zoological laboratories rarely exceed 1 m.
Subclass Elasmobranchii: Sharks, Skates, and Rays
There are nine living orders of elasmobranchs, numbering about 815 species. Coastal waters are dominated by the requiem sharks, order Carcharhiniformes, which consist of typical-looking sharks such as the tiger and bull sharks and more bizarre forms, including the hammerheads (Figure 26-7). The order Lamniformes contains several large, pelagic sharks dangerous to humans, including the great white and mako sharks. Dogfish sharks, familiar to generations of comparative anatomy students, are in the order Squaliformes. Skates and several groups of rays (sawfish rays, electric rays, stingrays, eagle rays, manta rays, and devil rays) belong to the order Rajiformes.
Much has been written about the propensities of sharks to attack humans, both by those exaggerating their ferocious nature and by those seeking to write them off as harmless. It is true, as the latter group of writers argues, that sharks are by nature timid and cautious. But it also is a fact that certain of them are dangerous to humans. There are numerous authenticated cases of shark attacks by Carcharodon (Gr. karcharos, sharp, + odous, tooth), the great white shark (reaching 6 m); mako sharks Isurus (Gr. is, equal, + ouros, tail); the tiger shark Galeocerdo (Gr. galeos, shark, + kerdo, fox); and hammerhead sharks Sphyrna (Gr. sphyra, hammer). More shark casualties have been reported from the tropical and temperate waters of the Australian region than from any other. During World War II there were several reports of mass shark attacks on the victims of ship sinkings in tropical waters.
Form and Function
Although to most people sharks have a sinister appearance and fearsome reputation, they are at the same time among the most gracefully streamlined of all fishes. The body of a dogfish shark (Figure 26-8) is fusiform (spindle shaped). In front of the ventral mouth is a pointed rostrum; at the posterior end the vertebral column turns up to end in the longer upper lobe of the tail. This type of tail is called heterocercal. The fins consist of paired pectoral and pelvic fins supported by appendicular skeletons, one or two median dorsal fins (each with a spine in Squalus [L. a kind of sea fish]), and a median caudal fin. A median anal fin is present in most sharks, including the smooth dogfish Mustelus (L. mustela, weasel). In the male, the medial part of the pelvic fin is modified to form a clasper, which is used in copulation. Paired nostrils (blind pouches) are ventral and anterior to the mouth (Figure 26-9). The lateral eyes are lidless, and behind each eye is a spiracle (remnant of the first gill slit). Five gill slits are found anterior to each pectoral fin. The tough, leathery skin is covered with toothlike, dermal placoid scales arranged to reduce the turbulence of water flowing along the body surface during swimming.
Sharks are well equipped for their predatory life. They track their prey using highly sensitive senses in an orderly sequence. Sharks may initially detect prey from a kilometer or more away with their large olfactory organs, capable of detecting chemicals as low as 1 part per 10 billion. The laterally placed nostrils of hammerhead sharks (Figure 26-7) may enhance odor localization by improving stereo-olfaction. Prey also may be located from long distances by sensing low-frequency vibrations with mechanoreceptors in the lateral line system. This system is composed of special receptor organs (neuromasts) in interconnected tubes and pores extending along the sides of the body and over the head (Figure 26- 10). At closer range the shark switches to vision as the primary method of tracking prey. Contrary to popular belief, most sharks have excellent vision, even in dimly lit waters. During the final stage of attack, sharks are guided to their prey by the bioelectric fields that surround all animals. Electroreceptors, the ampullae of Lorenzini (Figure 26-9), are located primarily on the shark’s head. In addition, sharks may use electroreception to find prey buried in the sand.
Both the upper and lower jaws of sharks are provided with many sharp, triangular teeth. The front row of functional teeth on the edge of the jaw is backed by rows of developing teeth that replace worn teeth throughout the life of the shark (Figure 26-8 and 26-9). The mouth cavity opens into the large pharynx, which contains openings to the separate gill slits and spiracles. A short, wide esophagus runs to the J-shaped stomach. A liver and pancreas open into the short, straight intestine, which contains the spiral valve that slows passage of food and increases the absorptive surface (Figure 26-11). Attached to the short rectum is the rectal gland, unique to chondrichthyans, which secretes a colorless fluid containing a high concentration of sodium chloride. It assists the opisthonephric kidney in regulating the salt concentration of the blood. The chambers of the heart are arranged in tandem formation, and the flow pattern of the circulatory system is basically the same as that of other gillbreathing vertebrates (Figure 26-11).
All chondrichthyans have internal fertilization, but maternal support of the embryo is highly variable. Many elasmobranchs lay large, yolky eggs immediately after fertilization; these species are termed oviparous. Some oviparous sharks and rays deposit their eggs in a horny capsule called a “mermaid’s purse,” which often is provided with tendrils that wrap around the first firm object it contacts, much like the tendrils of grape vines. The embryo is nourished from the yolk for a prolonged period—6 to 9 months in some, as much as 2 years in one species— before hatching as a miniature replica of the adult. Many sharks, however, retain the embryos in the reproductive tract for prolonged periods. Some are ovoviviparous species, which retain the developing young in the uterus while they are nourished by the contents of their yolk sac until born. Still other species have true viviparous reproduction. In these, embryos receive nourishment from the maternal bloodstream through the placenta, or from nutritive secretions, “uterine milk,” produced by the mother. Some sharks (sand tigers) exhibit a grisly type of reproduction in which embryos receive additional nutrition by eating eggs and siblings. The evolution of prolonged retention of embryos by many elasmobranchs was an important innovation that contributed to the success of these fish. However, regardless of the form of maternal support, once the eggs are laid, or the young born, all parental care ends.
Marine elasmobranchs have developed an interesting solution to the physiological problem of living in a salty medium. To prevent water from being drawn out of the body osmotically, elasmobranchs retain nitrogenous compounds, especially urea and trimethylamine oxide, in the blood. These solutes, combined with the blood salts, raise the blood solute concentration to exceed slightly that of seawater, eliminating an osmotic inequality between their bodies and the surrounding seawater.
A little more than half of all elasmobranchs are rays, a group that includes skates, electric rays, sawfishes, stingrays, eagle rays, and manta rays. Most are specialized for bottom dwelling, with a dorsoventrally flattened body and greatly enlarged pectoral fins that are fused to the head and used like wings in swimming (Figure 26-12). The gill openings are on the underside of the head, but the large spiracles are on top. Water for breathing is taken in through these spiracles to prevent clogging the gills, for the mouth is often buried in sand. Their teeth are adapted for crushing their prey: molluscs, crustaceans, and an occasional small fish.
The stingrays have a slender and whiplike tail that is armed with one or more saw-edged spines with venom glands at the base. Wounds from the spines are excruciatingly painful, and may heal slowly and with complications. Electric rays are sluggish fish with large electric organs on each side of the head (Figure 26-13). Each organ is made up of numerous vertical stacks of disclike cells connected in parallel so that when all the cells are discharged simultaneously, a high-amperage current is produced that flows out into the surrounding water. The voltage produced is relatively low (50 volts) but the power output may be almost one kilowatt— quite sufficient to stun prey or discourage predators. Electric rays were used by the ancient Egyptians for a form of electrotherapy in the treatment of afflictions such as arthritis and gout.
Subclass Holocephali: Chimaeras
Members of the small subclass Holocephali, distinguished by such suggestive names as ratfish (Figure 26-14), rabbitfish, spookfish, and ghostfish, are remnants of a line that diverged from the earliest shark lineage which originated at least 360 million years ago (Devonian or Silurian periods of the Paleozoic). Fossil chimaeras (ky-meer´- uz) first occurred in the Carboniferous period, reached their zenith in the Cretaceous and early Tertiary periods (120 million to 50 million years ago), and have declined ever since. Today there are only about 31 species extant.
Anatomically the chimaeras have several features linking them to elasmobranchs, but possess a suite of unique characters, too. Instead of a toothed mouth, their jaws bear large flat plates. The upper jaw is completely fused to the cranium, a most unusual development in fishes. Their food is seaweed, molluscs, echinoderms, crustaceans, and fishes—a surprisingly mixed diet for such a specialized grinding dentition. Chimaeras are not commercial species and are seldom caught. Despite their grotesque shape, they are beautifully colored with a pearly iridescence.
There are nearly 850 living species in the class Chondrichthyes, an ancient, compact, and highly developed group. Although a much smaller and less diverse assemblage than the bony fishes, their impressive combination of well-developed sense organs, powerful jaws and swimming musculature, and predaceous habits ensures them a secure and lasting place in the aquatic community. One of their distinctive features is their cartilaginous skeleton. Although calcification may be extensive in their skeletons, bone is entirely absent throughout the class—a curious evolutionary feature, since the Chondrichthyes are derived from ancestors having well-developed bone. Almost all chondrichthyans are marine; only 28 species live primarily in fresh water.
With the exception of whales, sharks include the largest living vertebrates. The larger sharks may reach 12 m in length. Dogfish sharks so widely studied in zoological laboratories rarely exceed 1 m.
Subclass Elasmobranchii: Sharks, Skates, and Rays
There are nine living orders of elasmobranchs, numbering about 815 species. Coastal waters are dominated by the requiem sharks, order Carcharhiniformes, which consist of typical-looking sharks such as the tiger and bull sharks and more bizarre forms, including the hammerheads (Figure 26-7). The order Lamniformes contains several large, pelagic sharks dangerous to humans, including the great white and mako sharks. Dogfish sharks, familiar to generations of comparative anatomy students, are in the order Squaliformes. Skates and several groups of rays (sawfish rays, electric rays, stingrays, eagle rays, manta rays, and devil rays) belong to the order Rajiformes.
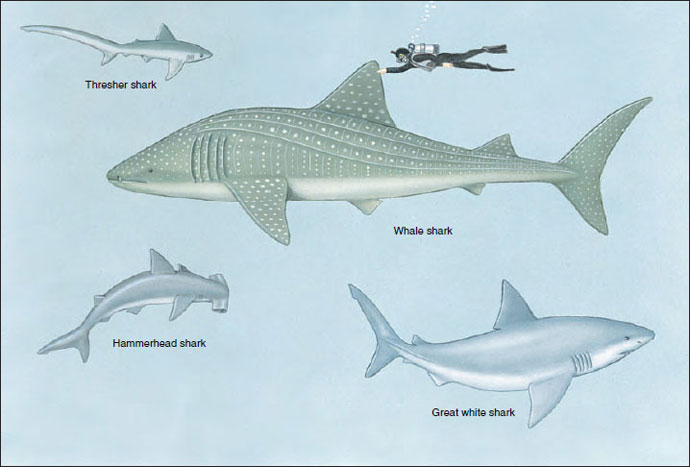 |
| Figure 26-7 Diversity in sharks of the subclass Elasmobranchii. The thresher shark Alopias vulpinus, exceptional because of its long upper tail lobe, may exceed 4 m in length. The great white shark Carcharodon carcharias, largest and most notorious of dangerous sharks, is a heavy-bodied, spindle-shaped shark that may reach 6 m in length. The nine species of hammerheads (genus Sphyrna) are distinguished from all other sharks by the flattened head with hammerlike lobes bearing eyes and nostrils on the ends. The whale shark Rhincodon typus is the world’s largest fish, reaching 12 m in length. It is a suspension feeder that feeds on plankton collected on a sievelike mesh over its gills. |
Much has been written about the propensities of sharks to attack humans, both by those exaggerating their ferocious nature and by those seeking to write them off as harmless. It is true, as the latter group of writers argues, that sharks are by nature timid and cautious. But it also is a fact that certain of them are dangerous to humans. There are numerous authenticated cases of shark attacks by Carcharodon (Gr. karcharos, sharp, + odous, tooth), the great white shark (reaching 6 m); mako sharks Isurus (Gr. is, equal, + ouros, tail); the tiger shark Galeocerdo (Gr. galeos, shark, + kerdo, fox); and hammerhead sharks Sphyrna (Gr. sphyra, hammer). More shark casualties have been reported from the tropical and temperate waters of the Australian region than from any other. During World War II there were several reports of mass shark attacks on the victims of ship sinkings in tropical waters.
Form and Function
Although to most people sharks have a sinister appearance and fearsome reputation, they are at the same time among the most gracefully streamlined of all fishes. The body of a dogfish shark (Figure 26-8) is fusiform (spindle shaped). In front of the ventral mouth is a pointed rostrum; at the posterior end the vertebral column turns up to end in the longer upper lobe of the tail. This type of tail is called heterocercal. The fins consist of paired pectoral and pelvic fins supported by appendicular skeletons, one or two median dorsal fins (each with a spine in Squalus [L. a kind of sea fish]), and a median caudal fin. A median anal fin is present in most sharks, including the smooth dogfish Mustelus (L. mustela, weasel). In the male, the medial part of the pelvic fin is modified to form a clasper, which is used in copulation. Paired nostrils (blind pouches) are ventral and anterior to the mouth (Figure 26-9). The lateral eyes are lidless, and behind each eye is a spiracle (remnant of the first gill slit). Five gill slits are found anterior to each pectoral fin. The tough, leathery skin is covered with toothlike, dermal placoid scales arranged to reduce the turbulence of water flowing along the body surface during swimming.
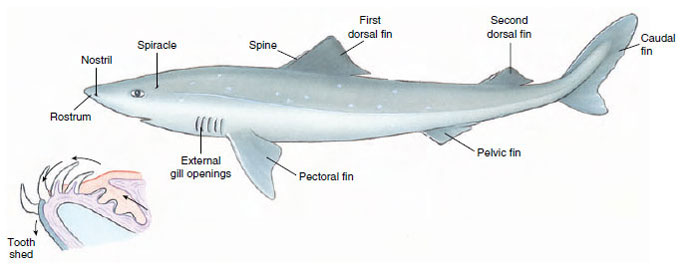 |
| Figure 26-8 Dogfish shark, Squalus acanthias. Inset: section of lower jaw shows new teeth developing inside the jaw. These move forward to replace lost teeth. Rate of replacement varies in different species. |
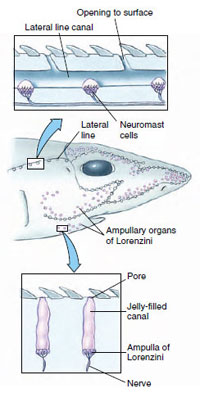 |
| Figure 26-10 Sensory canals and receptors in a shark. The ampullae of Lorenzini respond to weak electric fields, and possibly to temperature, water pressure, and salinity. The lateral line sensors, called neuromasts, are sensitive to disturbances in the water, enabling the shark to detect nearby objects by reflected waves in the water. |
 |
| Figure 26-9 Head of sand tiger shark Carcharias sp. Note the series of successional teeth. Also visible in a row below the eye are the ampullae of Lorenzini. |
Sharks are well equipped for their predatory life. They track their prey using highly sensitive senses in an orderly sequence. Sharks may initially detect prey from a kilometer or more away with their large olfactory organs, capable of detecting chemicals as low as 1 part per 10 billion. The laterally placed nostrils of hammerhead sharks (Figure 26-7) may enhance odor localization by improving stereo-olfaction. Prey also may be located from long distances by sensing low-frequency vibrations with mechanoreceptors in the lateral line system. This system is composed of special receptor organs (neuromasts) in interconnected tubes and pores extending along the sides of the body and over the head (Figure 26- 10). At closer range the shark switches to vision as the primary method of tracking prey. Contrary to popular belief, most sharks have excellent vision, even in dimly lit waters. During the final stage of attack, sharks are guided to their prey by the bioelectric fields that surround all animals. Electroreceptors, the ampullae of Lorenzini (Figure 26-9), are located primarily on the shark’s head. In addition, sharks may use electroreception to find prey buried in the sand.
Both the upper and lower jaws of sharks are provided with many sharp, triangular teeth. The front row of functional teeth on the edge of the jaw is backed by rows of developing teeth that replace worn teeth throughout the life of the shark (Figure 26-8 and 26-9). The mouth cavity opens into the large pharynx, which contains openings to the separate gill slits and spiracles. A short, wide esophagus runs to the J-shaped stomach. A liver and pancreas open into the short, straight intestine, which contains the spiral valve that slows passage of food and increases the absorptive surface (Figure 26-11). Attached to the short rectum is the rectal gland, unique to chondrichthyans, which secretes a colorless fluid containing a high concentration of sodium chloride. It assists the opisthonephric kidney in regulating the salt concentration of the blood. The chambers of the heart are arranged in tandem formation, and the flow pattern of the circulatory system is basically the same as that of other gillbreathing vertebrates (Figure 26-11).
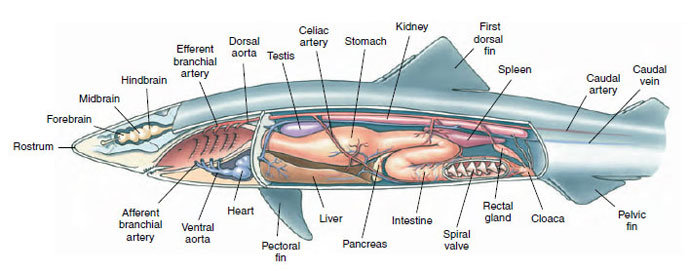 |
| Figure 26-11 Internal anatomy of dogfish shark Squalus acanthias. |
All chondrichthyans have internal fertilization, but maternal support of the embryo is highly variable. Many elasmobranchs lay large, yolky eggs immediately after fertilization; these species are termed oviparous. Some oviparous sharks and rays deposit their eggs in a horny capsule called a “mermaid’s purse,” which often is provided with tendrils that wrap around the first firm object it contacts, much like the tendrils of grape vines. The embryo is nourished from the yolk for a prolonged period—6 to 9 months in some, as much as 2 years in one species— before hatching as a miniature replica of the adult. Many sharks, however, retain the embryos in the reproductive tract for prolonged periods. Some are ovoviviparous species, which retain the developing young in the uterus while they are nourished by the contents of their yolk sac until born. Still other species have true viviparous reproduction. In these, embryos receive nourishment from the maternal bloodstream through the placenta, or from nutritive secretions, “uterine milk,” produced by the mother. Some sharks (sand tigers) exhibit a grisly type of reproduction in which embryos receive additional nutrition by eating eggs and siblings. The evolution of prolonged retention of embryos by many elasmobranchs was an important innovation that contributed to the success of these fish. However, regardless of the form of maternal support, once the eggs are laid, or the young born, all parental care ends.
Marine elasmobranchs have developed an interesting solution to the physiological problem of living in a salty medium. To prevent water from being drawn out of the body osmotically, elasmobranchs retain nitrogenous compounds, especially urea and trimethylamine oxide, in the blood. These solutes, combined with the blood salts, raise the blood solute concentration to exceed slightly that of seawater, eliminating an osmotic inequality between their bodies and the surrounding seawater.
A little more than half of all elasmobranchs are rays, a group that includes skates, electric rays, sawfishes, stingrays, eagle rays, and manta rays. Most are specialized for bottom dwelling, with a dorsoventrally flattened body and greatly enlarged pectoral fins that are fused to the head and used like wings in swimming (Figure 26-12). The gill openings are on the underside of the head, but the large spiracles are on top. Water for breathing is taken in through these spiracles to prevent clogging the gills, for the mouth is often buried in sand. Their teeth are adapted for crushing their prey: molluscs, crustaceans, and an occasional small fish.
 |
| Figure 26-12 Skates and rays are specialized for life on the sea floor. Both the clearnose skate Raja eglanteria (A), and the southern stingray Dasyatis americana (B) are flattened dorsoventrally and move by undulations of winglike pectoral fins. The stingray (B) is followed by a pilot fish. |
The stingrays have a slender and whiplike tail that is armed with one or more saw-edged spines with venom glands at the base. Wounds from the spines are excruciatingly painful, and may heal slowly and with complications. Electric rays are sluggish fish with large electric organs on each side of the head (Figure 26-13). Each organ is made up of numerous vertical stacks of disclike cells connected in parallel so that when all the cells are discharged simultaneously, a high-amperage current is produced that flows out into the surrounding water. The voltage produced is relatively low (50 volts) but the power output may be almost one kilowatt— quite sufficient to stun prey or discourage predators. Electric rays were used by the ancient Egyptians for a form of electrotherapy in the treatment of afflictions such as arthritis and gout.
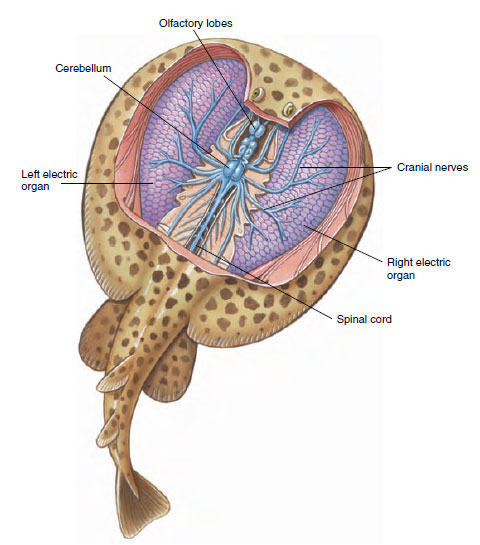 |
| Figure 26-13 Electric ray Torpedo with electric organs exposed. Organs are built up of disclike, multinucleated cells called electrocytes. When all cells are discharged simultaneously, a high-amperage current flows into the surrounding water to stun prey or discourage predators. Power output may be up to one kilowatt. |
 |
| Figure 26-14 Spotted ratfish, Hydrolagus collei, of North American west coast. This species is one of the most handsome of chimaeras, which tend toward bizarre appearances. |
Subclass Holocephali: Chimaeras
Members of the small subclass Holocephali, distinguished by such suggestive names as ratfish (Figure 26-14), rabbitfish, spookfish, and ghostfish, are remnants of a line that diverged from the earliest shark lineage which originated at least 360 million years ago (Devonian or Silurian periods of the Paleozoic). Fossil chimaeras (ky-meer´- uz) first occurred in the Carboniferous period, reached their zenith in the Cretaceous and early Tertiary periods (120 million to 50 million years ago), and have declined ever since. Today there are only about 31 species extant.
Anatomically the chimaeras have several features linking them to elasmobranchs, but possess a suite of unique characters, too. Instead of a toothed mouth, their jaws bear large flat plates. The upper jaw is completely fused to the cranium, a most unusual development in fishes. Their food is seaweed, molluscs, echinoderms, crustaceans, and fishes—a surprisingly mixed diet for such a specialized grinding dentition. Chimaeras are not commercial species and are seldom caught. Despite their grotesque shape, they are beautifully colored with a pearly iridescence.
Support our developers




