Class Oligochaeta
Class Oligochaeta
More than 3000 species of oligochaetes are found in a great variety of sizes and habitats. They include the familiar earthworms and many species that live in fresh water. Most are terrestrial or freshwater forms, but some are parasitic, and a few live in marine or brackish water.
With few exceptions, oligochaetes bear setae, which may be long or short, straight or curved, blunt or needlelike, or arranged singly or in bundles. Whatever the type, they are less numerous in oligochaetes than in polychaetes, as is implied by the class name, which means “few long hairs.” Aquatic forms usually have longer setae than do earthworms.
Earthworms
The most familiar of oligochaetes are earthworms (“night crawlers”), which burrow in moist, rich soil, emerging at night to explore their surroundings. In damp, rainy weather they stay near the surface, often with mouth or anus protruding from the burrow. In very dry weather they may burrow several feet underground, coil up in a slime chamber, and become dormant. Lumbricus terrestris (L. lumbricum, earthworm), the form commonly studied in school laboratories, is 12 to 30 cm long (Figure 17-12). Giant tropical earthworms may have from 150 to 250 or more segments and may grow to as much as 4 m in length. They usually live in branched and interconnected tunnels.
Form and Function
In earthworms the mouth is overhung by a fleshy prostomium at the anterior end, and the anus is on the posterior end (Figure 17-12B). In most earthworms each segment bears four pairs of chitinous setae (Figure 17-12C), although in some oligochaetes each segment may have up to 100 or more. Each seta is a bristlelike rod set in a sac within the body wall and moved by tiny muscles (Figure 17-13). The setae project through small pores in the cuticle to the outside. In locomotion and burrowing, setae anchor parts of the body to prevent slipping. Earthworms move by peristaltic movement. Contractions of circular muscles in the anterior end lengthen the body, pushing the anterior end forward where it is anchored by setae; contractions of longitudinal muscles then shorten the body, pulling the posterior end forward. As these waves of contraction pass along the entire body, it gradually moves forward.
Nutrition: Most oligochaetes are scavengers. Earthworms feed mainly on decayed organic matter, bits of leaves and vegetation, refuse, and animal matter. After being moistened by secretions from the mouth, food is drawn inward by the sucking action of the muscular pharynx. The liplike prostomium aids in manipulating food into position. Calcium from soil swallowed with food tends to produce a high blood calcium level. Calciferous glands along the esophagus secrete calcium ions into the gut and so reduce the calcium ion concentration of the blood. Calciferous glands are really ionoregulatory, rather than digestive, organs. They also function in regulating acid-base balance of body fluids, maintaining the pH at a fairly stable value.
Leaving the esophagus, food is stored temporarily in the thin-walled crop before being passed on into the gizzard, which grinds food into small pieces. Digestion and absorption occur in the intestine. Along the dorsal side, the wall of the intestine is infolded to form a typhlosole, which greatly increases the absorptive and digestive surface (Figure 17-12C). The digestive system secretes various enzymes to break down the food.
Surrounding the intestine and dorsal vessel and filling much of the typhlosole is a layer of yellowish chloragogen tissue derived from the peritoneum. This tissue serves as a center for synthesis of glycogen and fat, a function roughly equivalent to that of liver cells. Chloragogen cells when ripe (full of fat) are released into the coelom where they float free as cells called eleocytes (Gr. elaio, oil, + kytos, hollow vessel [cell]), which transport materials to the body tissues. They apparently can pass from segment to segment and have been found to accumulate around wounds and regenerating areas, where they break down and release their contents into the coelom. Chloragogen cells also function in excretion.
Circulation and Respiration: Annelids have a double transport system: coelomic fluid and circulatory system. Food, wastes, and respiratory gases are carried by both coelomic fluid and blood in varying degrees. Blood circulates in a closed system of vessels, including capillary systems in the tissues. Five main blood trunks run lengthwise through the body.
The dorsal vessel (single) runs above the alimentary canal from the pharynx to the anus. It is a pumping organ, provided with valves, and it functions as a true heart. This vessel receives blood from vessels of the body wall and digestive tract and pumps it anteriorly into five pairs of aortic arches. The function of aortic arches is to maintain a steady pressure of blood into the ventral vessel.
The ventral vessel (single) serves as the aorta. It receives blood from the aortic arches and delivers it to the brain and rest of the body, providing segmental vessels to the walls, nephridia, and digestive tract.
Their blood contains colorless ameboid cells and a dissolved respiratory pigment, hemoglobin. The blood of some annelids may have respiratory pigments other than hemoglobin, as noted above.
Earthworms have no special respiratory organs, but gaseous exchange takes place across their moist skin.
Excretion: Each somite except the first three and the last one bears a pair of metanephridia. Each nephridium occupies parts of two successive somites (Figure 17-14). A ciliated funnel, the nephrostome, lies just anterior to an intersegmental septum and leads by a small ciliated tubule through the septum into the somite behind, where it connects with the main part of the nephridium. Several complex loops of increasing size compose the nephridial duct, which terminates in a bladderlike structure leading to an aperture, the nephridiopore. The nephridiopore opens to the outside near the ventral row of setae. By means of cilia, wastes from the coelom are drawn into the nephrostome and tubule, where they are joined by salts and organic wastes transported from blood capillaries in the glandular part of the nephridium. Waste is discharged to the outside through the nephridiopore.
Aquatic oligochaetes excrete ammonia; terrestrial oligochaetes excrete the much less toxic urea. Lumbricus produces both, the level of urea depending somewhat on environmental conditions. Both urea and ammonia are produced by chloragogen cells, which may break off and enter nephridia directly, or their products may be carried by the blood. Some nitrogenous waste is eliminated through the body surface.
Oligochaetes are largely freshwater animals, and even such terrestrial forms as earthworms must exist in a moist environment. Osmoregulation is a function of the body surface and the nephridia, as well as the gut and dorsal pores. Lumbricus will gain weight when placed in tap water and lose it when returned to soil. Salts as well as water can pass across the integument, salts apparently being carried by active transport.
Nervous System and Sense Organs
The nervous system in earthworms (Figure 17-15) consists of a central system and peripheral nerves. The central system reflects the typical annelid pattern: a pair of cerebral ganglia (the brain) above the pharynx, a pair of connectives passing around the pharynx connecting the brain with the first pair of ganglia in the nerve cord; a ventral nerve cord, really double, running along the floor of the coelom to the last somite; and a pair of fused ganglia on the nerve cord in each somite. Each pair of fused ganglia provides nerves to the body structures, which contain both sensory and motor fibers.
Neurosecretory cells have been found in the brain and ganglia of annelids, both oligochaetes and polychaetes. They are endocrine in function and secrete neurohormones concerned with regulation of reproduction, secondary sex characteristics, and regeneration.
For rapid escape movements most annelids have from one to several very large axons commonly called giant axons (Figure 17-16), or giant fibers, located in the ventral nerve cord. Their large diameter increases rate of conduction and makes possible simultaneous contractions of muscles in many segments.
Simple sense organs are distributed all over the body. Earthworms have no eyes but do have many lens-shaped photoreceptors in their epidermis. Most oligochaetes are negatively phototactic to strong light but posi-tively phototactic to weak light. Many single-celled sense organs are widely distributed in the epidermis. What are presumably chemoreceptors are most numerous on the prostomium. There are many free nerve endings in the integument, which are probably tactile.
General Behavior: Earthworms are among the most defenseless of creatures, yet their abundance and wide distribution indicate their ability to survive. Although they have no specialized sense organs, they are sensitive to many stimuli. They react positively to mechanical stimuli when such stimuli are moderate and negatively to a strong vibration (such as footfall near them), which causes them to retire quickly into their burrows. They react to light, which they avoid unless it is very weak. Chemical responses aid them in the choice of food.
Chemical as well as tactile responses are very important to earthworms. They not only must sample the organic content of the soil to find food, but also must sense its texture, acidity, and calcium content.
Experiments show that earthworms have some learning ability. They can be taught to avoid an electric shock, and thus an association reflex can be produced in them. Darwin credited earthworms with a great deal of intelligence in pulling leaves into their burrows: he observed that they seized the leaves by the narrow end, the easiest way for drawing a leafshaped object into a small hole. Darwin assumed that seizure of leaves by worms did not result from random handling or from chance but was purposeful in its mechanism. However, investigations since Darwin’s time have shown that the process is mainly one of trial and error, for earthworms often seize a leaf several times before getting it right.
Reproduction and Development
Earthworms are monoecious (hermaphroditic); both male and female organs are found in the same animal (Figure 17-12A). In Lumbricus reproductive systems are found in somites 9 to 15. Two pairs of small testes and two pairs of sperm funnels are surrounded by three pairs of large seminal vesicles. Immature sperm from the testes mature in seminal vesicles, then pass into sperm funnels and down sperm ducts to the male genital pores in somite 15, where they are expelled during copulation. Eggs are discharged by a pair of small ovaries into the coelomic cavity, where ciliated funnels of the oviducts carry them outside through female genital pores on somite 14. Two pairs of seminal receptacles in somites 9 and 10 receive and store sperm from the mate during copulation.
Reproduction in earthworms may occur at any season, but they usually copulate at night during warm, moist weather (Figure 17-17). When mating, worms extend their anterior ends from their burrows and bring their ventral surfaces together (Figure 17-18). They are held together by mucus secreted by their clitellum (L. clitellae, packsaddle) and by special ventral setae, which penetrate each other’s bodies in the regions of contact. After discharge, sperm travel to seminal receptacles of the other worm in its seminal grooves. After copulation each worm secretes first a mucous tube and then a tough, chitinlike band that forms a cocoon around its clitellum. As the cocoon passes forward, eggs from the oviducts, albumin from skin glands, and sperm from the mate (stored in the seminal receptacles) pour into it. Fertilization of eggs then takes place within the cocoon. When the cocoon leaves the worm, its ends close, producing a lemon-shaped body. Embryogenesis occurs within the cocoon, and the form that hatches from the egg is a young worm similar to the adult. Thus development is direct with no metamorphosis. Juveniles do not develop a clitellum until they are sexually mature.
Freshwater Oligochaetes
Freshwater oligochaetes usually are smaller and have more conspicuous setae than do earthworms. They are more mobile than earthworms and tend to have better-developed sense organs. They are generally benthic forms that creep about on the substrate or burrow in soft mud. Aquatic oligochaetes are an important food source for fishes. A few are ectoparasitic.
Some of the more common freshwater oligochaetes are the 1 mm long Aeolosoma (Gr. aiolos, quick-moving, + soma, body) (Figure 17-19B), which contains red or green pigments, has bundles of setae, and is often found in hay cultures; the 2 to 4 mm long Nais (L. nais, water nymph), which is brownish and has two bundles of setae on anterior segments and four bundles of setae on each posterior segment; the 10 to 25 mm long Stylaria (Gr. stylos, pillar) (Figure 17-19A), with setae arranged like those of Nais, a prostomium extended into a long process, and black eyespots; the 5 to 10 mm long Dero, (Gr. dere, neck or throat), which is reddish, lives in tubes, and usually has 3 to 4 pairs of tail gills (Figure 17-19D); the 30 to 40 mm long Tubifex (L. tubus, tube, + faciens, to make or do) (Figure 17-19C), which is reddish and lives with its head in mud at the bottom of ponds and its tail waving in the water; the 10 to 15 mm long Chaetogaster (N.L. chaeta, bristle, + gastrula, belly), which has only ventral bundles of setae; and Enchytraeus (Gr. enchytraeus, living in an earthen pot), small whitish worms that live both in moist soil and in water. Some oligochaetes, such as Aeolosoma, may form chains of zooids asexually by transverse fission (Figure 17-19B).
More than 3000 species of oligochaetes are found in a great variety of sizes and habitats. They include the familiar earthworms and many species that live in fresh water. Most are terrestrial or freshwater forms, but some are parasitic, and a few live in marine or brackish water.
With few exceptions, oligochaetes bear setae, which may be long or short, straight or curved, blunt or needlelike, or arranged singly or in bundles. Whatever the type, they are less numerous in oligochaetes than in polychaetes, as is implied by the class name, which means “few long hairs.” Aquatic forms usually have longer setae than do earthworms.
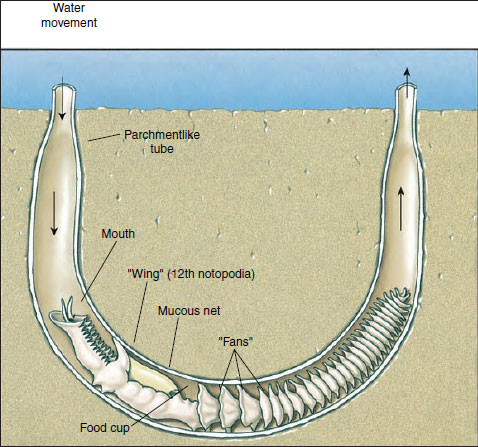 |
| Figure 17-11 Chaetopterus, a sedentary polychaete, lives in a U-shaped tube in the sea bottom. It pumps water through the parchmentlike tube (of which one-half has been cut away here) with its three pistonlike fans. The fans beat 60 times per minute to keep water currents moving. The winglike notopodia of the twelfth segment continuously secrete a mucous net that strains out food particles. As the net fills with food, the food cup rolls it into a ball, and when the ball is large enough (about 3 mm), the food cup bends forward and deposits the ball in a ciliated groove to be carried to the mouth and swallowed. |
Earthworms
The most familiar of oligochaetes are earthworms (“night crawlers”), which burrow in moist, rich soil, emerging at night to explore their surroundings. In damp, rainy weather they stay near the surface, often with mouth or anus protruding from the burrow. In very dry weather they may burrow several feet underground, coil up in a slime chamber, and become dormant. Lumbricus terrestris (L. lumbricum, earthworm), the form commonly studied in school laboratories, is 12 to 30 cm long (Figure 17-12). Giant tropical earthworms may have from 150 to 250 or more segments and may grow to as much as 4 m in length. They usually live in branched and interconnected tunnels.
 |
| Figure 17-12 Earthworm anatomy. A, Internal structure of anterior portion of worm. B, External features, lateral view. C, Generalized transverse section through region posterior to clitellum. D, Portion of epidermis showing sensory, glandular, and epithelial cells. |
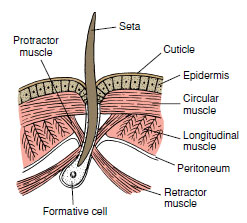 |
| Figure 17-13 Seta with its muscle attachments showing relation to adjacent structures. Setae lost by wear are replaced by new ones, which develop from formative cells. |
Form and Function
In earthworms the mouth is overhung by a fleshy prostomium at the anterior end, and the anus is on the posterior end (Figure 17-12B). In most earthworms each segment bears four pairs of chitinous setae (Figure 17-12C), although in some oligochaetes each segment may have up to 100 or more. Each seta is a bristlelike rod set in a sac within the body wall and moved by tiny muscles (Figure 17-13). The setae project through small pores in the cuticle to the outside. In locomotion and burrowing, setae anchor parts of the body to prevent slipping. Earthworms move by peristaltic movement. Contractions of circular muscles in the anterior end lengthen the body, pushing the anterior end forward where it is anchored by setae; contractions of longitudinal muscles then shorten the body, pulling the posterior end forward. As these waves of contraction pass along the entire body, it gradually moves forward.
Nutrition: Most oligochaetes are scavengers. Earthworms feed mainly on decayed organic matter, bits of leaves and vegetation, refuse, and animal matter. After being moistened by secretions from the mouth, food is drawn inward by the sucking action of the muscular pharynx. The liplike prostomium aids in manipulating food into position. Calcium from soil swallowed with food tends to produce a high blood calcium level. Calciferous glands along the esophagus secrete calcium ions into the gut and so reduce the calcium ion concentration of the blood. Calciferous glands are really ionoregulatory, rather than digestive, organs. They also function in regulating acid-base balance of body fluids, maintaining the pH at a fairly stable value.
Leaving the esophagus, food is stored temporarily in the thin-walled crop before being passed on into the gizzard, which grinds food into small pieces. Digestion and absorption occur in the intestine. Along the dorsal side, the wall of the intestine is infolded to form a typhlosole, which greatly increases the absorptive and digestive surface (Figure 17-12C). The digestive system secretes various enzymes to break down the food.
Surrounding the intestine and dorsal vessel and filling much of the typhlosole is a layer of yellowish chloragogen tissue derived from the peritoneum. This tissue serves as a center for synthesis of glycogen and fat, a function roughly equivalent to that of liver cells. Chloragogen cells when ripe (full of fat) are released into the coelom where they float free as cells called eleocytes (Gr. elaio, oil, + kytos, hollow vessel [cell]), which transport materials to the body tissues. They apparently can pass from segment to segment and have been found to accumulate around wounds and regenerating areas, where they break down and release their contents into the coelom. Chloragogen cells also function in excretion.
Circulation and Respiration: Annelids have a double transport system: coelomic fluid and circulatory system. Food, wastes, and respiratory gases are carried by both coelomic fluid and blood in varying degrees. Blood circulates in a closed system of vessels, including capillary systems in the tissues. Five main blood trunks run lengthwise through the body.
The dorsal vessel (single) runs above the alimentary canal from the pharynx to the anus. It is a pumping organ, provided with valves, and it functions as a true heart. This vessel receives blood from vessels of the body wall and digestive tract and pumps it anteriorly into five pairs of aortic arches. The function of aortic arches is to maintain a steady pressure of blood into the ventral vessel.
The ventral vessel (single) serves as the aorta. It receives blood from the aortic arches and delivers it to the brain and rest of the body, providing segmental vessels to the walls, nephridia, and digestive tract.
Their blood contains colorless ameboid cells and a dissolved respiratory pigment, hemoglobin. The blood of some annelids may have respiratory pigments other than hemoglobin, as noted above.
Earthworms have no special respiratory organs, but gaseous exchange takes place across their moist skin.
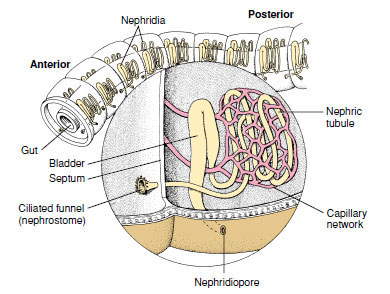 |
| Figure 17-14 Nephridium of earthworm. Wastes are drawn into the ciliated nephrostome in one segment, then passed through the loops of the nephridium, and expelled through the nephridiopore of the next segment. |
Excretion: Each somite except the first three and the last one bears a pair of metanephridia. Each nephridium occupies parts of two successive somites (Figure 17-14). A ciliated funnel, the nephrostome, lies just anterior to an intersegmental septum and leads by a small ciliated tubule through the septum into the somite behind, where it connects with the main part of the nephridium. Several complex loops of increasing size compose the nephridial duct, which terminates in a bladderlike structure leading to an aperture, the nephridiopore. The nephridiopore opens to the outside near the ventral row of setae. By means of cilia, wastes from the coelom are drawn into the nephrostome and tubule, where they are joined by salts and organic wastes transported from blood capillaries in the glandular part of the nephridium. Waste is discharged to the outside through the nephridiopore.
Aquatic oligochaetes excrete ammonia; terrestrial oligochaetes excrete the much less toxic urea. Lumbricus produces both, the level of urea depending somewhat on environmental conditions. Both urea and ammonia are produced by chloragogen cells, which may break off and enter nephridia directly, or their products may be carried by the blood. Some nitrogenous waste is eliminated through the body surface.
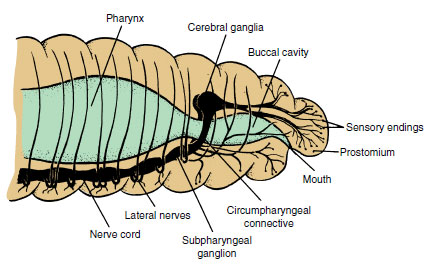 |
| Figure 17-15 Anterior portion of earthworm and its nervous system. Note concentration of sensory endings in this region. |
Oligochaetes are largely freshwater animals, and even such terrestrial forms as earthworms must exist in a moist environment. Osmoregulation is a function of the body surface and the nephridia, as well as the gut and dorsal pores. Lumbricus will gain weight when placed in tap water and lose it when returned to soil. Salts as well as water can pass across the integument, salts apparently being carried by active transport.
Nervous System and Sense Organs
The nervous system in earthworms (Figure 17-15) consists of a central system and peripheral nerves. The central system reflects the typical annelid pattern: a pair of cerebral ganglia (the brain) above the pharynx, a pair of connectives passing around the pharynx connecting the brain with the first pair of ganglia in the nerve cord; a ventral nerve cord, really double, running along the floor of the coelom to the last somite; and a pair of fused ganglia on the nerve cord in each somite. Each pair of fused ganglia provides nerves to the body structures, which contain both sensory and motor fibers.
Neurosecretory cells have been found in the brain and ganglia of annelids, both oligochaetes and polychaetes. They are endocrine in function and secrete neurohormones concerned with regulation of reproduction, secondary sex characteristics, and regeneration.
For rapid escape movements most annelids have from one to several very large axons commonly called giant axons (Figure 17-16), or giant fibers, located in the ventral nerve cord. Their large diameter increases rate of conduction and makes possible simultaneous contractions of muscles in many segments.
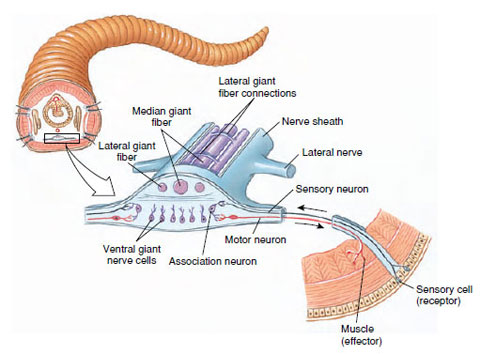 |
| Figure 17-16 Portion of nerve cord of earthworm showing arrangement of simple reflex arc (in foreground) and the three dorsal giant fibers that are adapted for rapid reflexes and escape movements. Ordinary crawling involves a succession of reflex acts, the stretching of one somite stimulating the next to stretch. Impulses are transmitted much faster in giant fibers than in regular nerves so that all segments can contract simultaneously when quick withdrawal into a burrow is necessary. |
Simple sense organs are distributed all over the body. Earthworms have no eyes but do have many lens-shaped photoreceptors in their epidermis. Most oligochaetes are negatively phototactic to strong light but posi-tively phototactic to weak light. Many single-celled sense organs are widely distributed in the epidermis. What are presumably chemoreceptors are most numerous on the prostomium. There are many free nerve endings in the integument, which are probably tactile.
General Behavior: Earthworms are among the most defenseless of creatures, yet their abundance and wide distribution indicate their ability to survive. Although they have no specialized sense organs, they are sensitive to many stimuli. They react positively to mechanical stimuli when such stimuli are moderate and negatively to a strong vibration (such as footfall near them), which causes them to retire quickly into their burrows. They react to light, which they avoid unless it is very weak. Chemical responses aid them in the choice of food.
Chemical as well as tactile responses are very important to earthworms. They not only must sample the organic content of the soil to find food, but also must sense its texture, acidity, and calcium content.
Experiments show that earthworms have some learning ability. They can be taught to avoid an electric shock, and thus an association reflex can be produced in them. Darwin credited earthworms with a great deal of intelligence in pulling leaves into their burrows: he observed that they seized the leaves by the narrow end, the easiest way for drawing a leafshaped object into a small hole. Darwin assumed that seizure of leaves by worms did not result from random handling or from chance but was purposeful in its mechanism. However, investigations since Darwin’s time have shown that the process is mainly one of trial and error, for earthworms often seize a leaf several times before getting it right.
 |
| Figure 17-17 Two earthworms in copulation. Their anterior ends point in opposite directions as their ventral surfaces are held together by mucous bands secreted by the clitella. Mutual insemination occurs during copulation. After separation each worm secretes a cocoon to receive its eggs and sperm. |
Reproduction and Development
Earthworms are monoecious (hermaphroditic); both male and female organs are found in the same animal (Figure 17-12A). In Lumbricus reproductive systems are found in somites 9 to 15. Two pairs of small testes and two pairs of sperm funnels are surrounded by three pairs of large seminal vesicles. Immature sperm from the testes mature in seminal vesicles, then pass into sperm funnels and down sperm ducts to the male genital pores in somite 15, where they are expelled during copulation. Eggs are discharged by a pair of small ovaries into the coelomic cavity, where ciliated funnels of the oviducts carry them outside through female genital pores on somite 14. Two pairs of seminal receptacles in somites 9 and 10 receive and store sperm from the mate during copulation.
Reproduction in earthworms may occur at any season, but they usually copulate at night during warm, moist weather (Figure 17-17). When mating, worms extend their anterior ends from their burrows and bring their ventral surfaces together (Figure 17-18). They are held together by mucus secreted by their clitellum (L. clitellae, packsaddle) and by special ventral setae, which penetrate each other’s bodies in the regions of contact. After discharge, sperm travel to seminal receptacles of the other worm in its seminal grooves. After copulation each worm secretes first a mucous tube and then a tough, chitinlike band that forms a cocoon around its clitellum. As the cocoon passes forward, eggs from the oviducts, albumin from skin glands, and sperm from the mate (stored in the seminal receptacles) pour into it. Fertilization of eggs then takes place within the cocoon. When the cocoon leaves the worm, its ends close, producing a lemon-shaped body. Embryogenesis occurs within the cocoon, and the form that hatches from the egg is a young worm similar to the adult. Thus development is direct with no metamorphosis. Juveniles do not develop a clitellum until they are sexually mature.
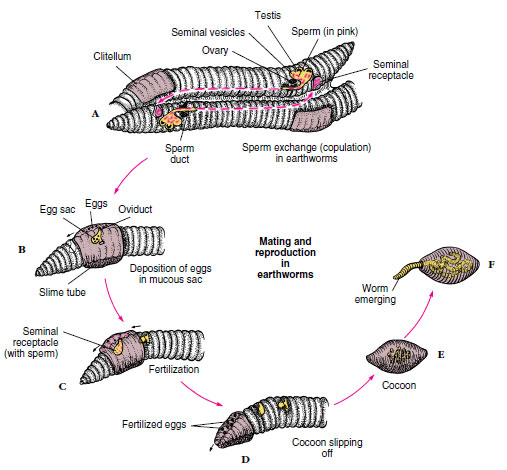 |
| Figure 17-18 Earthworm copulation and formation of egg cocoons. A, Mutual insemination; sperm from genital pore (somite 15) pass along seminal grooves to seminal receptacles (somites 9 and 10) of each mate. B and C, After worms separate, a slime tube formed over the clitellum passes forward to receive eggs from oviducts and sperm from seminal receptacles. D, As cocoon slips off over anterior end, its ends close and seal. E, Cocoon is deposited near burrow entrance. F, Young worms emerge in 2 to 3 weeks. |
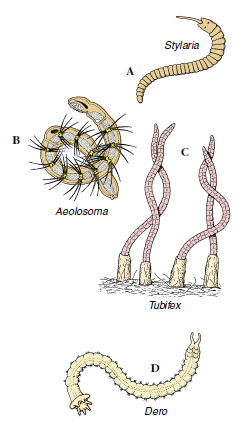 |
| Figure 17-19 Some freshwater oligochaetes. A, Stylaria has the prostomium drawn out into a long snout. B, Aeolosoma uses cilia around the mouth to sweep in food particles, and it buds off new individuals asexually. C, Tubifex lives head down in long tubes. D, Dero has ciliated anal gills. |
Freshwater Oligochaetes
Freshwater oligochaetes usually are smaller and have more conspicuous setae than do earthworms. They are more mobile than earthworms and tend to have better-developed sense organs. They are generally benthic forms that creep about on the substrate or burrow in soft mud. Aquatic oligochaetes are an important food source for fishes. A few are ectoparasitic.
Some of the more common freshwater oligochaetes are the 1 mm long Aeolosoma (Gr. aiolos, quick-moving, + soma, body) (Figure 17-19B), which contains red or green pigments, has bundles of setae, and is often found in hay cultures; the 2 to 4 mm long Nais (L. nais, water nymph), which is brownish and has two bundles of setae on anterior segments and four bundles of setae on each posterior segment; the 10 to 25 mm long Stylaria (Gr. stylos, pillar) (Figure 17-19A), with setae arranged like those of Nais, a prostomium extended into a long process, and black eyespots; the 5 to 10 mm long Dero, (Gr. dere, neck or throat), which is reddish, lives in tubes, and usually has 3 to 4 pairs of tail gills (Figure 17-19D); the 30 to 40 mm long Tubifex (L. tubus, tube, + faciens, to make or do) (Figure 17-19C), which is reddish and lives with its head in mud at the bottom of ponds and its tail waving in the water; the 10 to 15 mm long Chaetogaster (N.L. chaeta, bristle, + gastrula, belly), which has only ventral bundles of setae; and Enchytraeus (Gr. enchytraeus, living in an earthen pot), small whitish worms that live both in moist soil and in water. Some oligochaetes, such as Aeolosoma, may form chains of zooids asexually by transverse fission (Figure 17-19B).
Support our developers




