Class Polychaeta
Class Polychaeta
Polychaetes form the largest class of annelids with more than 10,000 species, most of them marine. Although most are 5 to 10 cm long, some are less than 1 mm, and others may be as long as 3 m. Some are brightly colored in reds and greens; others are dull or iridescent. Some are picturesque, such as the “featherduster” worms (Figure 17-2).
Polychaetes differ from other annelids in having a well-differentiated head with specialized sense organs; paired appendages, called parapodia, on most segments; and no clitellum (Figure 17-3). As their name implies, they have many setae, usually arranged in bundles on the parapodia. They show a pronounced differentiation of some body somites and a specialization of sensory organs practically unknown among clitellates.
Many polychaetes are euryhaline (can tolerate a wide range of environmental salinity) and occur in brackish water. The freshwater polychaete fauna is more diversified in warmer regions than in the temperate zones.
Polychaetes live under rocks, in coral crevices, or in abandoned shells, or they burrow into mud or sand; some build their own tubes on submerged objects or in bottom material; some adopt the tubes or homes of other animals; some are pelagic, making up a part of the planktonic population. They are extremely abundant in some areas; for example, a square meter of mudflat may contain thousands of polychaetes. They play a significant part in marine food chains because they are eaten by fish, crustaceans, hydroids, and many others.
They are often divided for convenience into two groups (formerly the basis of subclasses): sedentary polychaetes, and errant or free-moving polychaetes. Sedentary polychaetes are mainly tubicolous, spending all or much of their time in tubes or permanent burrows. Many of them, especially those that live in tubes, have elaborate devices for feeding and respiration. Errant polychaetes (L. errare, to wander), include free-moving pelagic forms, active burrowers, crawlers, and tube worms that leave their tubes for feeding or breeding. Most of these, like clam worms Nereis (Gr. name of a sea nymph) (Figure 17- 3), are predatory and equipped with jaws or teeth. They have a muscular eversible pharynx armed with teeth that can be thrust out with surprising speed and dexterity for capturing prey.
Form and Function
The polychaete typically has a head, or prostomium, which may or may not be retractile and which often bears eyes, tentacles, and sensory palps (Figures 17-3 and 17-7). The first segment (peristomium) surrounds the mouth and may bear setae, palps, or, in predatory forms, chitinous jaws. Ciliary feeders may bear a tentacular crown that can be opened like a fan or withdrawn into the tube.
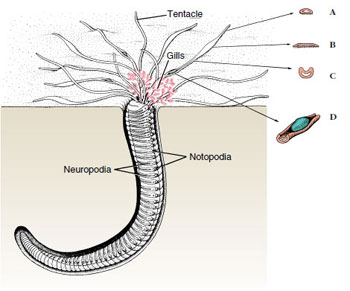
The trunk is segmented, and most segments bear parapodia, which may have lobes, cirri, setae, and other parts on them (see Figure 17-3). Parapodia are used in crawling, swimming, or anchoring in tubes. They usually serve as the chief respiratory organs, although some polychaetes also have gills. Amphitrite, for example, has three pairs of branched gills and long extensible tentacles (Figure 17-4). Arenicola, the lugworm (Figure 17-5), which burrows through sand leaving characteristic castings at the entrance to its burrow, has paired gills on certain somites.
Nutrition
The polychaete digestive system consists of a foregut, midgut, and hindgut. The foregut includes a stomodeum, pharynx, and anterior esophagus. It is lined with cuticle, and the jaws, where present, are constructed of cuticular protein. The midgut is derived from endoderm. More anterior portions secrete digestive enzymes, and absorption takes place toward the posterior. The short, ectodermally-derived hindgut connects the midgut to the exterior via the anus, which is on the pygidium.
Errant polychaetes are mostly predators and scavengers. Sedentary polychaetes feed on suspended particles, or they are deposit feeders, consuming particles on or in the sediment. We discuss food habits of some specific polychaetes in the following text.
Circulation and Respiration
Polychaetes show considerable diversity in both circulatory and respiratory structure and function. As mentioned before, parapodia and gills serve for gaseous exchange in various species. In some polychaetes there are no special organs for respiration, and gaseous exchange takes place across the body surface.
The circulatory pattern varies greatly. In Nereis a dorsal longitudinal vessel carries blood anteriorly, and a ventral longitudinal vessel conducts it posteriorly (Figure 17-3C). Blood flows between these two vessels via segmental networks in the parapodia, septa, and around the intestine. In Glycera the circulatory system is reduced and joins with the coelom. Septa are incomplete, and thus the coelomic fluid assumes the function of circulation.
Many polychaetes have respiratory pigments such as hemoglobin, chlorocruorin, or hemerythrin.
Excretion
Although there is some variety in excretory organs, including possession of protonephridia and mixed proto- and metanephridia in some, most polychaetes have metanephridia (Figure 17-3). There is one pair per metamere, with the inner end of each (nephrostome) opening into a coelomic compartment. Coelomic fluid passes into the nephrostome, and selective resorption occurs along the nephridial duct, as in oligochaetes (see Figure 17-14).
Nervous System and Sense Organs
Organization of the central nervous system in polychaetes follows the basic annelid plan (see Figure 17-15). Dorsal cerebral ganglia connect with a subpharyngeal ganglion via a circumpharyngeal commissure. A double ventral nerve cord courses the length of the worm, with metamerically arranged ganglia.
Sense organs are more highly developed in polychaetes than in oligochaetes and include eyes, nuchal organs, and statocysts. Eyes, when present, may range from simple eyespots to well-developed organs. Eyes are most conspicuous in errant worms. Usually the eyes are retinal cups, with rodlike photoreceptor cells lining the cup wall and directed toward the lumen of the cup. The highest degree of development occurs in the family Alciopidae, which has large, imageresolving eyes similar in structure to those of some cephalopod molluscs (Figure 16-39), with cornea, lens, retina, and retinal pigment. Alciopid eyes also have accessory retinas, a characteristic shared by deepsea fishes and some deep-sea cephalopods. Different wavelengths of light penetrate to different depths in water, and the accessory retinas of alciopids are sensitive to different wavelengths. The eyes of these pelagic animals may be well adapted to function as the light varies with depth. Studies with electroencephalograms show that they are sensitive to dim light of the deep sea. Nuchal organs are ciliated sensory pits or slits that appear to be chemoreceptive, an important factor in food gathering. Some burrowing and tubebuilding polychaetes have statocysts that function in body orientation.
Reproduction and Development
In contrast to clitellates, polychaetes have no permanent sex organs, and they usually have separate sexes. Reproductive systems are simple. Gonads appear as temporary swellings of the peritoneum and shed their gametes into the coelom. They are carried outside through gonoducts, through the metanephridia, or by rupture of the body wall. Fertilization is external, and the early larva is a trochophore (see Figure 16-6).
Clam Worms: Nereis
Clam worms (Figure 17-7), or sand worms as they are sometimes called, are errant polychaetes that live in mucus-lined burrows in or near low tide. Sometimes they are found in temporary hiding places, such as under stones, where they stay with their bodies covered and their heads protruding. They are most active at night, when they wiggle out of their hiding places and swim about or crawl over the sand in search of food.
The body, containing about 200 somites, may grow to 30 or 40 cm in length. The head is made up of a prostomium and a peristomium. The prostomium bears a pair of stubby palps, sensitive to touch and taste; a pair of short sensory tentacles; and two pairs of small dorsal eyes that are light sensitive. The peristomium bears the ventral mouth, a pair of chitinous jaws, and four pairs of sensory tentacles (Figure 17-3A).
Each parapodium has two lobes: a dorsal notopodium and a ventral neuropodium (Figure 17-3C). One or more chitinous spines (acicula) supports each lobe. Parapodia bear setae and are abundantly supplied with blood vessels. Parapodia are used for both creeping and swimming and are manipulated by oblique muscles that run from the midventral line to the parapodia in each somite. The worm swims by lateral undulatory wriggling of the body—unlike the peristaltic movement of earthworms. It can dart through the water with considerable speed. These undulatory movements can also be used to suck water into or pump it out of the burrow. The worm will usually adapt some kind of burrow if it can find one. When a worm is placed near a glass tube, it will wriggle in without hesitation.
Clam worms feed on small animals, other worms, and larval forms. They seize food with their chitinous jaws, which they protrude through the mouth when they evert their pharynx. As they withdraw the pharynx, they swallow the food. Movement of food through the alimentary canal is by peristalsis.
Other Interesting Polychaetes
Scale worms (Figure 17-8) are members of the family Polynoidae (Gr. Polynoe, daughter of Nereus and Doris, a sea god and goddess), one of the most abundant and widespread of polychaete families. Their rather flattened bodies are covered with broad scales, modified from dorsal parts of parapodia. Most are of modest size, but some are enormous (up to 190 mm long and 100 mm wide). They are carnivorous and feed on a wide variety of animals. Many are commensal, living in burrows of other polychaetes or in association with cnidarians, molluscs, or echinoderms.
Hermodice carunculata (Gr. herma, reef, + dex, a worm found in wood) (Figure 17-9) and related species are called fireworms. Their setae are hollow, brittle, and contain a poisonous secretion. When touched, the setae break off in the wound and cause skin irritation. They feed on corals, gorgonians, and other cnidarians.
Tube dwellers secrete many types of tubes. Some are parchmentlike or leathery (Figure 17-2B); some are firm, calcareous tubes attached to rocks or other surfaces (Figure 17-2A); and some are simply grains of sand or bits of shell or seaweed cemented together with mucous secretions. Many burrowers in sand and mud flats simply line their burrows with mucus (Figure 17-5).
Most sedentary tube and burrow dwellers are particle feeders, using ciliary or mucoid methods of obtaining food. The principal food source is plankton and detritus. Some, like Amphitrite (Gr. a mythical sea nymph) (Figure 17-4), with head peeping out of the mud, send out long extensible tentacles over the surface to deposit feed. Cilia and mucus on the tentacles entrap particles found on the sea bottom and move them toward the mouth. Lugworms Arenicola (L. arena, sand, + colo, inhabit) employ an interesting combination of suspension and deposit feeding. They live in an Lshaped burrow in which, by peristaltic movements, they cause water to flow. Food particles are filtered out by the sand at the front of the burrow, and they ingest the food-laden sand (Figure 17-5).
17-5). Fanworms, or “featherduster” worms, are beautiful tubeworms, fascinating to watch as they emerge from their secreted tubes and unfurl their lovely tentacular crowns to feed (Figure 17-2). A slight disturbance, sometimes even a passing shadow, causes them to duck quickly into the safety of the homes they have built. Food attracted to the feathery arms, or radioles, by ciliary action is trapped in mucus and is carried down ciliated food grooves to the mouth (Figure 17-10). Particles too large for the food grooves pass along the margins and drop off. Further sorting may occur near the mouth where only small particles of food enter the mouth, and sand grains are stored in a sac to be used later in enlarging the tube.
The parchment worm Chaetopterus (Gr. chaite, long hair, + pteron, wing) feeds on suspended particles by an entirely different mechanism (Figure 17-11). It lives in a U-shaped, parchmentlike tube buried, except for the tapered ends, in sand or mud along the shore. The worm attaches to the side of the tube by ventral suckers. Fans (modified parapodia) on segments 14 to 16 pump water through the tube by rhythmical movements. A pair of enlarged parapodia in the twelfth segment secretes a long mucous bag that reaches back to a small food cup just in front of the fans. All water passing through the tube is filtered through this mucous bag, the end of which is rolled into a ball by cilia in the cup. When the ball is about the size of a BB shot, the fans stop beating and the ball of food and mucus is rolled forward by ciliary action to the mouth and swallowed.
Polychaetes form the largest class of annelids with more than 10,000 species, most of them marine. Although most are 5 to 10 cm long, some are less than 1 mm, and others may be as long as 3 m. Some are brightly colored in reds and greens; others are dull or iridescent. Some are picturesque, such as the “featherduster” worms (Figure 17-2).
 |
| Figure 17-2 Tube-dwelling sedentary polychaetes. A, One of the featherduster worms (called a Christmas-tree worm), Spirobranchus giganteus, has a double crown of radioles and lives in a calcareous tube. B, Sabellid polychaetes, Bispira brunnea, live in leathery tubes. |
Polychaetes differ from other annelids in having a well-differentiated head with specialized sense organs; paired appendages, called parapodia, on most segments; and no clitellum (Figure 17-3). As their name implies, they have many setae, usually arranged in bundles on the parapodia. They show a pronounced differentiation of some body somites and a specialization of sensory organs practically unknown among clitellates.
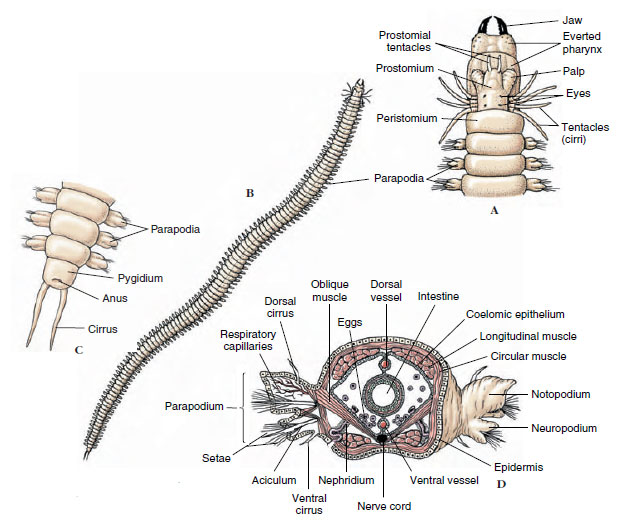 |
| Figure 17-3 Nereis virens, an errant polychaete. A, Anterior end, with pharynx everted. B, External structure. C, Posterior end. D, Generalized transverse section through region of the intestine. |
Many polychaetes are euryhaline (can tolerate a wide range of environmental salinity) and occur in brackish water. The freshwater polychaete fauna is more diversified in warmer regions than in the temperate zones.
 |
| Figure 17-4 Amphitrite, which builds its tubes in mud or sand, extends long grooved tentacles out over the mud to pick up bits of organic matter. The smallest particles are moved along food grooves by cilia, larger particles by peristaltic movement. Its plumelike gills are blood red. A, Section through exploratory end of tentacle. B, Section through tentacle in area adhering to substratum. C, Section showing ciliary groove. D, Particle being carried toward mouth. |
Polychaetes live under rocks, in coral crevices, or in abandoned shells, or they burrow into mud or sand; some build their own tubes on submerged objects or in bottom material; some adopt the tubes or homes of other animals; some are pelagic, making up a part of the planktonic population. They are extremely abundant in some areas; for example, a square meter of mudflat may contain thousands of polychaetes. They play a significant part in marine food chains because they are eaten by fish, crustaceans, hydroids, and many others.
They are often divided for convenience into two groups (formerly the basis of subclasses): sedentary polychaetes, and errant or free-moving polychaetes. Sedentary polychaetes are mainly tubicolous, spending all or much of their time in tubes or permanent burrows. Many of them, especially those that live in tubes, have elaborate devices for feeding and respiration. Errant polychaetes (L. errare, to wander), include free-moving pelagic forms, active burrowers, crawlers, and tube worms that leave their tubes for feeding or breeding. Most of these, like clam worms Nereis (Gr. name of a sea nymph) (Figure 17- 3), are predatory and equipped with jaws or teeth. They have a muscular eversible pharynx armed with teeth that can be thrust out with surprising speed and dexterity for capturing prey.
Form and Function
The polychaete typically has a head, or prostomium, which may or may not be retractile and which often bears eyes, tentacles, and sensory palps (Figures 17-3 and 17-7). The first segment (peristomium) surrounds the mouth and may bear setae, palps, or, in predatory forms, chitinous jaws. Ciliary feeders may bear a tentacular crown that can be opened like a fan or withdrawn into the tube.
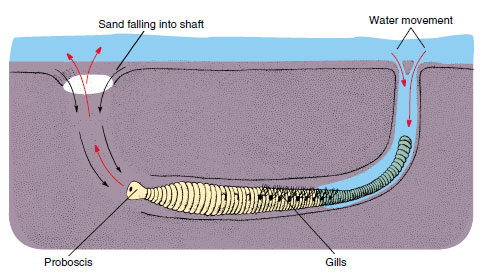 |
| Figure 17-5 Arenicola, the lugworm, lives in an L-shaped burrow in intertidal mudflats. It burrows by successive eversions and retractions of its proboscis. By peristaltic movements it keeps water filtering through the sand. The worm then ingests the food-laden sand. |
The trunk is segmented, and most segments bear parapodia, which may have lobes, cirri, setae, and other parts on them (see Figure 17-3). Parapodia are used in crawling, swimming, or anchoring in tubes. They usually serve as the chief respiratory organs, although some polychaetes also have gills. Amphitrite, for example, has three pairs of branched gills and long extensible tentacles (Figure 17-4). Arenicola, the lugworm (Figure 17-5), which burrows through sand leaving characteristic castings at the entrance to its burrow, has paired gills on certain somites.
Nutrition
The polychaete digestive system consists of a foregut, midgut, and hindgut. The foregut includes a stomodeum, pharynx, and anterior esophagus. It is lined with cuticle, and the jaws, where present, are constructed of cuticular protein. The midgut is derived from endoderm. More anterior portions secrete digestive enzymes, and absorption takes place toward the posterior. The short, ectodermally-derived hindgut connects the midgut to the exterior via the anus, which is on the pygidium.
Errant polychaetes are mostly predators and scavengers. Sedentary polychaetes feed on suspended particles, or they are deposit feeders, consuming particles on or in the sediment. We discuss food habits of some specific polychaetes in the following text.
Circulation and Respiration
Polychaetes show considerable diversity in both circulatory and respiratory structure and function. As mentioned before, parapodia and gills serve for gaseous exchange in various species. In some polychaetes there are no special organs for respiration, and gaseous exchange takes place across the body surface.
The circulatory pattern varies greatly. In Nereis a dorsal longitudinal vessel carries blood anteriorly, and a ventral longitudinal vessel conducts it posteriorly (Figure 17-3C). Blood flows between these two vessels via segmental networks in the parapodia, septa, and around the intestine. In Glycera the circulatory system is reduced and joins with the coelom. Septa are incomplete, and thus the coelomic fluid assumes the function of circulation.
Many polychaetes have respiratory pigments such as hemoglobin, chlorocruorin, or hemerythrin.
Excretion
Although there is some variety in excretory organs, including possession of protonephridia and mixed proto- and metanephridia in some, most polychaetes have metanephridia (Figure 17-3). There is one pair per metamere, with the inner end of each (nephrostome) opening into a coelomic compartment. Coelomic fluid passes into the nephrostome, and selective resorption occurs along the nephridial duct, as in oligochaetes (see Figure 17-14).
Nervous System and Sense Organs
Organization of the central nervous system in polychaetes follows the basic annelid plan (see Figure 17-15). Dorsal cerebral ganglia connect with a subpharyngeal ganglion via a circumpharyngeal commissure. A double ventral nerve cord courses the length of the worm, with metamerically arranged ganglia.
Sense organs are more highly developed in polychaetes than in oligochaetes and include eyes, nuchal organs, and statocysts. Eyes, when present, may range from simple eyespots to well-developed organs. Eyes are most conspicuous in errant worms. Usually the eyes are retinal cups, with rodlike photoreceptor cells lining the cup wall and directed toward the lumen of the cup. The highest degree of development occurs in the family Alciopidae, which has large, imageresolving eyes similar in structure to those of some cephalopod molluscs (Figure 16-39), with cornea, lens, retina, and retinal pigment. Alciopid eyes also have accessory retinas, a characteristic shared by deepsea fishes and some deep-sea cephalopods. Different wavelengths of light penetrate to different depths in water, and the accessory retinas of alciopids are sensitive to different wavelengths. The eyes of these pelagic animals may be well adapted to function as the light varies with depth. Studies with electroencephalograms show that they are sensitive to dim light of the deep sea. Nuchal organs are ciliated sensory pits or slits that appear to be chemoreceptive, an important factor in food gathering. Some burrowing and tubebuilding polychaetes have statocysts that function in body orientation.
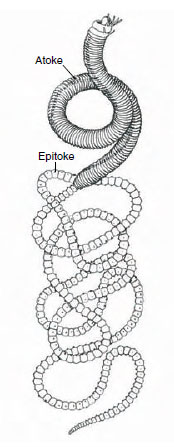 |
| Figure 17-6 Eunice viridis, the Samoan palolo worm. The posterior segments make up the epitokal region, consisting of segments packed with gametes. Each segment has an eyespot on the ventral side. Once a year the worms swarm, and the epitokes detach, rise to the surface, and discharge their ripe gametes, leaving the water milky. By the next breeding season, the epitokes are regenerated. |
Reproduction and Development
In contrast to clitellates, polychaetes have no permanent sex organs, and they usually have separate sexes. Reproductive systems are simple. Gonads appear as temporary swellings of the peritoneum and shed their gametes into the coelom. They are carried outside through gonoducts, through the metanephridia, or by rupture of the body wall. Fertilization is external, and the early larva is a trochophore (see Figure 16-6).
Clam Worms: Nereis
Clam worms (Figure 17-7), or sand worms as they are sometimes called, are errant polychaetes that live in mucus-lined burrows in or near low tide. Sometimes they are found in temporary hiding places, such as under stones, where they stay with their bodies covered and their heads protruding. They are most active at night, when they wiggle out of their hiding places and swim about or crawl over the sand in search of food.
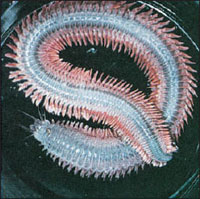 |
| Figure 17-7 Nereis diversicolor. Note the well-defined segments, the lobed parapodia, and the prostomium with tentacles. |
The body, containing about 200 somites, may grow to 30 or 40 cm in length. The head is made up of a prostomium and a peristomium. The prostomium bears a pair of stubby palps, sensitive to touch and taste; a pair of short sensory tentacles; and two pairs of small dorsal eyes that are light sensitive. The peristomium bears the ventral mouth, a pair of chitinous jaws, and four pairs of sensory tentacles (Figure 17-3A).
Each parapodium has two lobes: a dorsal notopodium and a ventral neuropodium (Figure 17-3C). One or more chitinous spines (acicula) supports each lobe. Parapodia bear setae and are abundantly supplied with blood vessels. Parapodia are used for both creeping and swimming and are manipulated by oblique muscles that run from the midventral line to the parapodia in each somite. The worm swims by lateral undulatory wriggling of the body—unlike the peristaltic movement of earthworms. It can dart through the water with considerable speed. These undulatory movements can also be used to suck water into or pump it out of the burrow. The worm will usually adapt some kind of burrow if it can find one. When a worm is placed near a glass tube, it will wriggle in without hesitation.
Clam worms feed on small animals, other worms, and larval forms. They seize food with their chitinous jaws, which they protrude through the mouth when they evert their pharynx. As they withdraw the pharynx, they swallow the food. Movement of food through the alimentary canal is by peristalsis.
Other Interesting Polychaetes
Scale worms (Figure 17-8) are members of the family Polynoidae (Gr. Polynoe, daughter of Nereus and Doris, a sea god and goddess), one of the most abundant and widespread of polychaete families. Their rather flattened bodies are covered with broad scales, modified from dorsal parts of parapodia. Most are of modest size, but some are enormous (up to 190 mm long and 100 mm wide). They are carnivorous and feed on a wide variety of animals. Many are commensal, living in burrows of other polychaetes or in association with cnidarians, molluscs, or echinoderms.
 |
| Figure 17-8 A scale worm Hesperonoe adventor normally lives as a commensal in the tubes of Urechis (Phylum Echiura) |
Hermodice carunculata (Gr. herma, reef, + dex, a worm found in wood) (Figure 17-9) and related species are called fireworms. Their setae are hollow, brittle, and contain a poisonous secretion. When touched, the setae break off in the wound and cause skin irritation. They feed on corals, gorgonians, and other cnidarians.
 |
| Figure 17-9 A fireworm Hermodice carunculata feeds on gorgonians and stony corals. Its setae are like tiny glass fibers and serve to ward off predators. |
Tube dwellers secrete many types of tubes. Some are parchmentlike or leathery (Figure 17-2B); some are firm, calcareous tubes attached to rocks or other surfaces (Figure 17-2A); and some are simply grains of sand or bits of shell or seaweed cemented together with mucous secretions. Many burrowers in sand and mud flats simply line their burrows with mucus (Figure 17-5).
Most sedentary tube and burrow dwellers are particle feeders, using ciliary or mucoid methods of obtaining food. The principal food source is plankton and detritus. Some, like Amphitrite (Gr. a mythical sea nymph) (Figure 17-4), with head peeping out of the mud, send out long extensible tentacles over the surface to deposit feed. Cilia and mucus on the tentacles entrap particles found on the sea bottom and move them toward the mouth. Lugworms Arenicola (L. arena, sand, + colo, inhabit) employ an interesting combination of suspension and deposit feeding. They live in an Lshaped burrow in which, by peristaltic movements, they cause water to flow. Food particles are filtered out by the sand at the front of the burrow, and they ingest the food-laden sand (Figure 17-5).
17-5). Fanworms, or “featherduster” worms, are beautiful tubeworms, fascinating to watch as they emerge from their secreted tubes and unfurl their lovely tentacular crowns to feed (Figure 17-2). A slight disturbance, sometimes even a passing shadow, causes them to duck quickly into the safety of the homes they have built. Food attracted to the feathery arms, or radioles, by ciliary action is trapped in mucus and is carried down ciliated food grooves to the mouth (Figure 17-10). Particles too large for the food grooves pass along the margins and drop off. Further sorting may occur near the mouth where only small particles of food enter the mouth, and sand grains are stored in a sac to be used later in enlarging the tube.
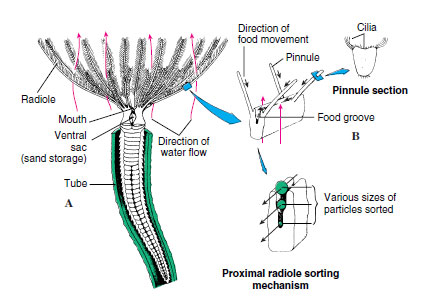 |
| Figure 17-10 Sabella, a polychaete ciliary feeder, extends its crown of feeding radioles from its leathery secreted tube, reinforced with sand and debris. A, Anterior view of the crown. Cilia direct small food particles along grooved radioles to mouth and discard larger particles. Sand grains are directed to storage sacs and later are used in tube building. B, Distal portion of radiole showing ciliary tracts of pinnules and food grooves. |
The parchment worm Chaetopterus (Gr. chaite, long hair, + pteron, wing) feeds on suspended particles by an entirely different mechanism (Figure 17-11). It lives in a U-shaped, parchmentlike tube buried, except for the tapered ends, in sand or mud along the shore. The worm attaches to the side of the tube by ventral suckers. Fans (modified parapodia) on segments 14 to 16 pump water through the tube by rhythmical movements. A pair of enlarged parapodia in the twelfth segment secretes a long mucous bag that reaches back to a small food cup just in front of the fans. All water passing through the tube is filtered through this mucous bag, the end of which is rolled into a ball by cilia in the cup. When the ball is about the size of a BB shot, the fans stop beating and the ball of food and mucus is rolled forward by ciliary action to the mouth and swallowed.
Support our developers




