Early Evolution of Terrestrial Vertebrates
Early Evolution
of Terrestrial
Vertebrates
Devonian Origin of Tetrapods
The Devonian period, beginning some 400 million years ago, was a time of mild temperatures and alternating droughts and floods. During this period, some primarily aquatic vertebrates evolved two features that would be important for permitting the subsequent evolution for life on land: lungs and limbs.
The Devonian freshwater environment was unstable. During dry periods, many pools and streams evaporated, water became foul, and dissolved oxygen disappeared. Only those fishes able to acquire atmospheric oxygen survived such conditions. Gills were unsuitable because in air the filaments collapsed, dried, and quickly lost their function. Virtually all freshwater fishes surviving this period, including lobefinned fishes and lungfishes, had a kind of lung that developed as an outgrowth of the pharynx. The efficiency of the air-filled cavity was enhanced by improving its vascularity with a rich capillary network, and by supplying it with arterial blood from the last (sixth) pair of aortic arches. Oxygenated blood returned directly to the heart by a pulmonary vein to form a complete pulmonary circuit. Thus the double circulation characteristic of all tetrapods originated: a systemic circulation serving the body and a pulmonary circulation supplying the lungs.
Vertebrate limbs also arose during the Devonian period. Although fish fins at first appear very different from the jointed limbs of tetrapods, an examination of the bony elements of the paired fins of lobe-finned fishes shows that they broadly resemble the equivalent limbs of amphibians. In Eusthenopteron, a Devonian lobe-fin, we can recognize an upper arm bone (humerus) and two forearm bones (radius and ulna) as well as other elements that we can homologize with the wrist bones of tetrapods (Figure 27-1). Eusthenopteron could walk—more accurately flop—along the bottom mud of pools with its fins, since backward and forward movement of the fins was limited to about 20 to 25 degrees. Acanthostega, one of the earliest known Devonian tetrapods, had well-formed tetrapod legs with clearly formed digits on both foreand hindlimbs, but the limbs were too weakly constructed to enable the animal to hoist its body off the surface for proper walking on land. Ichthyostega, however, with a fully developed shoulder girdle, bulky limb bones, welldeveloped muscles, and other adaptations for terrestrial life, must have been able to pull itself onto land, although it is doubtful that it could have walked very well.
Until recently zoologists thought that the early tetrapods had five fingers and five toes on their hands and feet, the basic pentadactyl plan of most living tetrapods today. However, newly discovered fossils of Devonian tetrapods show that all of them had more than five digits. Only later did the five-digit pattern become stabilized in the different tetrapod lineages.
Movement onto land was clearly a revolution in vertebrate history. How did it occur? A long-accepted scenario developed by Harvard paleontologist Alfred Romer suggested that when freshwater pools of the Devonian evaporated during seasonal droughts, the aquatic vertebrates were forced to move to others that still contained water. The fleshy fins of the sarcopterygians could be adapted as paddles to lever their way across land in search of water. Those with strong fins lived to reproduce. According to this hypothesis, land travel and the gradual development of legs originated as a means for survival in water. Recent discovery of more complete fossils of the earliest known tetrapods changes this view. Although Acanthostega had tetrapod legs (Figure 27-1), in every other respect it was a fully aquatic animal. A consensus emerging now is that tetrapods evolved their legs underwater and only then, for reasons unknown, began to pull themselves onto land.
As noted above, evidence points to lobe-finned fishes as the closest relatives of tetrapods; in cladistic terms they contain the sister group of tetrapods (Figures 27-2 and 27-3). Both lobe-finned fishes and early tetrapods such as Acanthostega and Ichthyostega shared several characteristics of skull, teeth, and pectoral girdle. Ichthyostega (Gr. ichthys, fish, + stege, roof, or covering, in reference to the roof of the skull, which was shaped like that of a fish) represents an early offshoot of tetrapod phylogeny that possessed several adaptations, in addition to jointed limbs, that equipped it for life on land. These include a stronger backbone and associated muscles to support the body in air, new muscles to elevate the head, strengthened shoulder and hip girdles, a protective rib cage, a more advanced ear structure for detecting airborne sounds, a foreshortening of the skull, and a lengthening of the snout that improved olfactory powers for detecting dilute airborne odors. Yet Ichthyostega still resembled aquatic forms in retaining a tail complete with fin rays and in having opercular (gill) bones.
Carboniferous Radiation of Tetrapods
The capricious Devonian period was followed by the Carboniferous period, characterized by a warm, wet climate during which mosses and large ferns grew in profusion on a swampy landscape. Tetrapods radiated quickly in this environment to produce a great variety of forms, feeding on the abundance of insects, insect larvae, and aquatic invertebrates available. Evolutionary relationships of early tetrapod groups are still controversial. We present a tentative cladogram (Figure 27-2), which almost certainly will undergo future revision as new data are collected. Several extinct lineages plus Lissamphibia, which contains modern amphibians, are placed in a group termed the temnospondyls. This group is distinguished by having generally only four digits on the forelimb rather than the five characteristic of most tetrapods.
Lissamphibians diversified during the Carboniferous to produce ancestors of the three major groups of amphibians alive today, frogs (Anura or Salientia), salamanders (Caudata or Urodela), and caecilians (Apoda or Gymnophiona). Amphibians improved their adaptations for living in water during this period. Their bodies became flatter for moving through shallow water. Early salamanders developed weak limbs and their tail became better developed as a swimming organ. Even anurans (frogs and toads), which are now largely terrestrial as adults, developed specialized hindlimbs with webbed feet better suited for swimming than for movement on land. All amphibians use their porous skin as a primary or accessory breathing organ. This specialization was encouraged by swampy environments of the Carboniferous period but presented serious desiccation problems for life on land.
Two additional generally recognized but nonetheless controversial groupings of Carboniferous and Permian tetrapods, lepospondyls and anthracosaurs, are judged on the basis of skull structure to be closer to amniotes than to temnospondyls (see Figure 27-3). Together they form a second major branch of tetrapod phylogeny that will be covered in Reptilian Groups , Birds and Mammals .
Devonian Origin of Tetrapods
The Devonian period, beginning some 400 million years ago, was a time of mild temperatures and alternating droughts and floods. During this period, some primarily aquatic vertebrates evolved two features that would be important for permitting the subsequent evolution for life on land: lungs and limbs.
The Devonian freshwater environment was unstable. During dry periods, many pools and streams evaporated, water became foul, and dissolved oxygen disappeared. Only those fishes able to acquire atmospheric oxygen survived such conditions. Gills were unsuitable because in air the filaments collapsed, dried, and quickly lost their function. Virtually all freshwater fishes surviving this period, including lobefinned fishes and lungfishes, had a kind of lung that developed as an outgrowth of the pharynx. The efficiency of the air-filled cavity was enhanced by improving its vascularity with a rich capillary network, and by supplying it with arterial blood from the last (sixth) pair of aortic arches. Oxygenated blood returned directly to the heart by a pulmonary vein to form a complete pulmonary circuit. Thus the double circulation characteristic of all tetrapods originated: a systemic circulation serving the body and a pulmonary circulation supplying the lungs.
Vertebrate limbs also arose during the Devonian period. Although fish fins at first appear very different from the jointed limbs of tetrapods, an examination of the bony elements of the paired fins of lobe-finned fishes shows that they broadly resemble the equivalent limbs of amphibians. In Eusthenopteron, a Devonian lobe-fin, we can recognize an upper arm bone (humerus) and two forearm bones (radius and ulna) as well as other elements that we can homologize with the wrist bones of tetrapods (Figure 27-1). Eusthenopteron could walk—more accurately flop—along the bottom mud of pools with its fins, since backward and forward movement of the fins was limited to about 20 to 25 degrees. Acanthostega, one of the earliest known Devonian tetrapods, had well-formed tetrapod legs with clearly formed digits on both foreand hindlimbs, but the limbs were too weakly constructed to enable the animal to hoist its body off the surface for proper walking on land. Ichthyostega, however, with a fully developed shoulder girdle, bulky limb bones, welldeveloped muscles, and other adaptations for terrestrial life, must have been able to pull itself onto land, although it is doubtful that it could have walked very well.
Until recently zoologists thought that the early tetrapods had five fingers and five toes on their hands and feet, the basic pentadactyl plan of most living tetrapods today. However, newly discovered fossils of Devonian tetrapods show that all of them had more than five digits. Only later did the five-digit pattern become stabilized in the different tetrapod lineages.
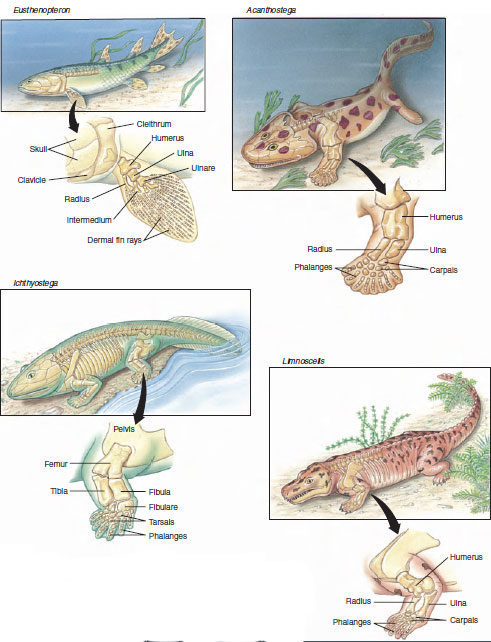 |
| Figure 27-1 Evolution of the tetrapod leg. Legs of tetrapods evolved from fins of Paleozoic fishes. Eusthenopteron, a late Devonian lobe-finned fish (rhipidistian) had paired muscular fins supported by bony elements that foreshadowed the bones of tetrapod limbs. The anterior fin contained an upper arm bone (humerus), two forerm bones (radius and ulna), and smaller elements homologous to wrist bones of tetrapods. As typical of fishes, the pectoral girdle, consisting of the cleithrum, clavicle, and other bones, was firmly attached to the skull. In Acanthostega, one of the earliest known Devonian tetrapods (appearing about 360 million years BP), dermal fin rays of the anterior appendage were replaced by eight fully evolved fingers. Acanthostega was probably exclusively aquatic because its limbs were too weak for travel on land. Ichthyostega, a contemporary of Acanthostega, had fully formed tetrapod limbs and must have been able to walk on land. The hindlimb bore seven toes (the number of front limb digits is unknown). Limnoscelis, an anthracosaur amphibian of the Carboniferous (about 300 million years BP) had five digits on both front and hindlimbs, the basic pentadactyl model that became the tetrapod standard. |
Movement onto land was clearly a revolution in vertebrate history. How did it occur? A long-accepted scenario developed by Harvard paleontologist Alfred Romer suggested that when freshwater pools of the Devonian evaporated during seasonal droughts, the aquatic vertebrates were forced to move to others that still contained water. The fleshy fins of the sarcopterygians could be adapted as paddles to lever their way across land in search of water. Those with strong fins lived to reproduce. According to this hypothesis, land travel and the gradual development of legs originated as a means for survival in water. Recent discovery of more complete fossils of the earliest known tetrapods changes this view. Although Acanthostega had tetrapod legs (Figure 27-1), in every other respect it was a fully aquatic animal. A consensus emerging now is that tetrapods evolved their legs underwater and only then, for reasons unknown, began to pull themselves onto land.
As noted above, evidence points to lobe-finned fishes as the closest relatives of tetrapods; in cladistic terms they contain the sister group of tetrapods (Figures 27-2 and 27-3). Both lobe-finned fishes and early tetrapods such as Acanthostega and Ichthyostega shared several characteristics of skull, teeth, and pectoral girdle. Ichthyostega (Gr. ichthys, fish, + stege, roof, or covering, in reference to the roof of the skull, which was shaped like that of a fish) represents an early offshoot of tetrapod phylogeny that possessed several adaptations, in addition to jointed limbs, that equipped it for life on land. These include a stronger backbone and associated muscles to support the body in air, new muscles to elevate the head, strengthened shoulder and hip girdles, a protective rib cage, a more advanced ear structure for detecting airborne sounds, a foreshortening of the skull, and a lengthening of the snout that improved olfactory powers for detecting dilute airborne odors. Yet Ichthyostega still resembled aquatic forms in retaining a tail complete with fin rays and in having opercular (gill) bones.
Carboniferous Radiation of Tetrapods
The capricious Devonian period was followed by the Carboniferous period, characterized by a warm, wet climate during which mosses and large ferns grew in profusion on a swampy landscape. Tetrapods radiated quickly in this environment to produce a great variety of forms, feeding on the abundance of insects, insect larvae, and aquatic invertebrates available. Evolutionary relationships of early tetrapod groups are still controversial. We present a tentative cladogram (Figure 27-2), which almost certainly will undergo future revision as new data are collected. Several extinct lineages plus Lissamphibia, which contains modern amphibians, are placed in a group termed the temnospondyls. This group is distinguished by having generally only four digits on the forelimb rather than the five characteristic of most tetrapods.
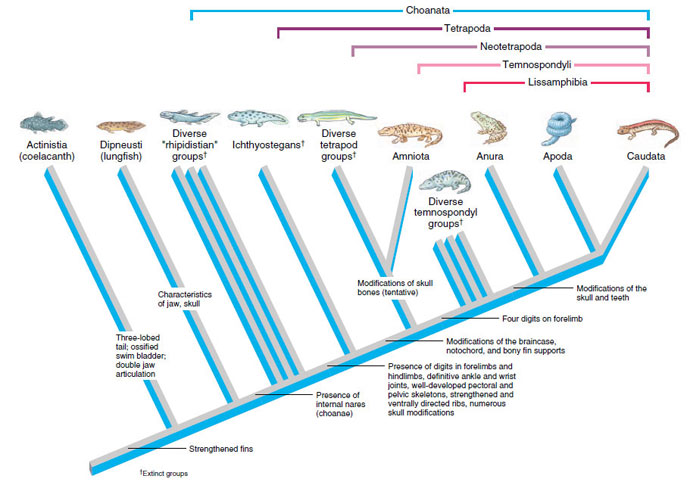 |
| Figure 27-2 Tentative cladogram of the Tetrapoda with emphasis on descent of amphibians. Especially controversial are relationships of major tetrapod groups (Amniota, Anthracosauria, Lepospondyli, Temnospondyli) and outgroups (Actinistia, Dipneusti, Rhipidistia). All aspects of this cladogram are controversial, however, including monophyly of Lissamphibia. Relationships shown for the three groups of Lissamphibia are based on recent molecular evidence. |
Lissamphibians diversified during the Carboniferous to produce ancestors of the three major groups of amphibians alive today, frogs (Anura or Salientia), salamanders (Caudata or Urodela), and caecilians (Apoda or Gymnophiona). Amphibians improved their adaptations for living in water during this period. Their bodies became flatter for moving through shallow water. Early salamanders developed weak limbs and their tail became better developed as a swimming organ. Even anurans (frogs and toads), which are now largely terrestrial as adults, developed specialized hindlimbs with webbed feet better suited for swimming than for movement on land. All amphibians use their porous skin as a primary or accessory breathing organ. This specialization was encouraged by swampy environments of the Carboniferous period but presented serious desiccation problems for life on land.
Two additional generally recognized but nonetheless controversial groupings of Carboniferous and Permian tetrapods, lepospondyls and anthracosaurs, are judged on the basis of skull structure to be closer to amniotes than to temnospondyls (see Figure 27-3). Together they form a second major branch of tetrapod phylogeny that will be covered in Reptilian Groups , Birds and Mammals .
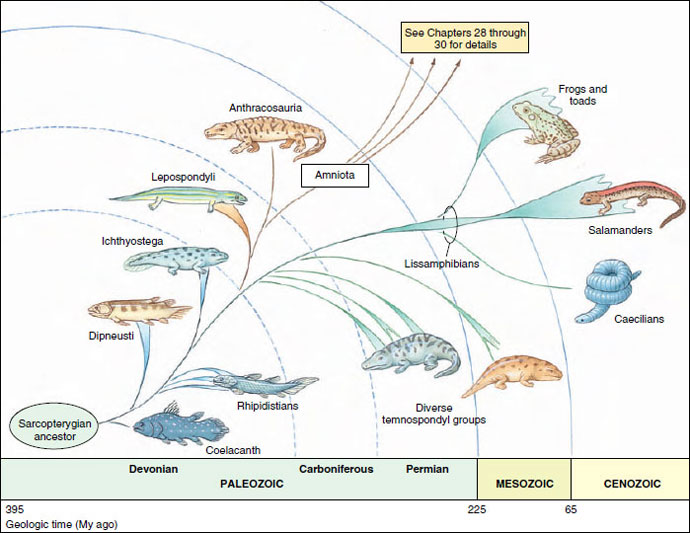 |
| Figure 27-3 Early tetrapod evolution and the descent of amphibians. Tetrapods share most recent common ancestry with Devonian rhipidistians. Amphibians share most recent common ancestry with diverse temnospondyls of the Carboniferous and Permian periods of the Paleozoic, and Triassic period of the Mesozoic. |
Support our developers




