Modern Amphibians
Modern Amphibians
The three living amphibian orders comprise more than 4200 species. Most share general adaptations for life on land, including skeletal strengthening and a shifting of special sense priorities from the ancestral lateral line system to the senses of smell and hearing. The olfactory epithelium and the ear are redesigned to improve sensitivities to airborne odors and sounds, respectively.
Nonetheless, most amphibians meet problems of independent life on land only halfway. In the ancestral life history of amphibians, eggs are aquatic and hatch to produce an aquatic larval form that uses gills for respiration. A metamorphosis follows in which gills are lost and lungs, which are present throughout larval life, are then activated for respiration. Many amphibians retain this general pattern but there are some important exceptions. Some salamanders lack a complete metamorphosis and retain a permanently aquatic, larval morphology throughout life. Others live entirely on land and lack the aquatic larval phase completely. Both of these are evolutionarily derived conditions. Some frogs also have acquired a strictly terrestrial existence by eliminating the aquatic larval stage. Some frogs, salamanders, and caecilians that undergo the complete metamorphic life cycle nonetheless remain in water as adults rather than moving onto land during their metamorphosis.
Even the most terrestrial amphibians remain dependent on very moist if not aquatic environments. Their skin is thin, and it requires moisture for protection against desiccation in air. An intact frog loses water nearly as rapidly as a skinless frog. Amphibians also require moderately cool environments. Being ectothermic, their body temperature is determined by and varies with the environment, greatly restricting where they can live. Cool and wet environments are especially important for reproduction. Eggs are not well protected from desiccation, and they must be shed directly into water or onto moist terrestrial surfaces. Completely terrestrial amphibians may lay eggs under logs or rocks, in the moist forest floor, in flooded tree holes, in pockets on the mother’s back (Figure 27-4), or in folds of the body wall. One species of Australian frog even broods its young in its vocal pouch.
We now highlight special characteristics of the three major groups of amphibians. We will expand coverage of general amphibian features when discussing groups in which particular features have been studied most extensively. For most features, this group will be the frogs.
Characteristics of Modern Amphibians
Caecilians: Order Gymnophiona (Apoda)
The order Gymnophiona (jim´no-fy´ona)
(Gr. gymnos, naked, + opineos, of a
snake) contains approximately 160
species of elongate, limbless, burrowing
creatures commonly called caecilians (Figure 27-5). They occur in tropical
forests of South America (their principal
home), Africa, and Southeast Asia. Caecilians
possess a long, slender body,
small scales in the skin of some, many
vertebrae, long ribs, no limbs, and a terminal
anus. Eyes are small, and most
species are totally blind as adults. Special
sensory tentacles occur on the
snout. Because they are almost entirely
burrowing or aquatic, they seldom are
seen by humans. Their food consists
mostly of worms and small invertebrates,
which they find underground.
Fertilization is internal, and the male is
provided with a protrusible copulatory
organ. Eggs usually are deposited in moist ground near water. Larvae may
be aquatic, or complete larval development
may occur in the egg. In some
species eggs are carefully guarded during
their development in folds of the
body. Viviparity also is common in
some caecilians, with embryos obtaining
nourishment by eating the wall of
the oviduct.
Salamanders: Order Caudata (Urodela)
As its name suggests, order Caudata (L. caudatus, having a tail) consists of tailed amphibians, approximately 360 species of salamanders. Salamanders are found in almost all northern temperate regions of the world, and they are abundant and diverse in North America. Salamanders are found also in tropical areas of Central and northern South America. Salamanders are typically small; most of the common North American salamanders are less than 15 cm long. Some aquatic forms are considerably longer, and the Japanese giant salamander may exceed 1.5 m in length.
Most salamanders have limbs set at right angles to the body, with forelimbs and hindlimbs of approximately equal size. In some aquatic and burrowing forms, limbs are rudimentary and some may be absent.
Salamanders are carnivorous both as larvae and adults, preying on worms, small arthropods, and small molluscs. Most eat only things that are moving. Since their food is rich in proteins, they do not store great quantities of fat or glycogen. Like all amphibians, they are ectotherms with a low metabolic rate.
Breeding Behavior
Some salamanders are aquatic throughout their life cycle, but most are metamorphic, having aquatic larvae and terrestrial adults that live in moist places under stones and rotten logs. Eggs of most salamanders are fertilized internally, usually after the female recovers in her vent a packet of sperm (spermatophore) that previously has been deposited by the male on a leaf or stick (Figure 27-6). Aquatic species lay their eggs in clusters or stringy masses in water. Their eggs hatch to produce an aquatic larva having external gills and a finlike tail. Completely terrestrial species deposit eggs in small, grapelike clusters under logs or in excavations in soft moist earth, and many species remain to guard the egggs (Figure 27-7). Terrestrial species have direct development: they bypass the larval stage and hatch as miniature versions of their parents. The most complex of salamander life cycles is observed in some American newts,
whose aquatic larvae metamorphose to
form terrestrial juveniles that later
metamorphose again to produce secondarily
aquatic, breeding adults (Figure
27-8). Many newt populations skip
the terrestrial “red eft” stage, however,
remaining entirely aquatic.
Respiration
Salamanders demonstrate an unusually diverse array of respiratory mechanisms. They share the general amphibian condition of having in their skin extensive vascular nets that serve respiratory exchange of oxygen and carbon dioxide. At various stages of their life history, salamanders also may have external gills, lungs, both, or neither of these. Salamanders with an aquatic larval stage hatch with gills, but lose them later if a metamorphosis occurs. Several diverse lineages of salamanders have evolved permanently aquatic forms that fail to undergo a complete metamorphosis and retain their gills and finlike tail throughout life. Lungs, the most widespread respiratory organ of terrestrial vertebrates, are present from birth in salamanders that have them, and become active following metamorphosis.
Although we normally associate lungs with terrestrial organisms and gills with aquatic ones, salamander evolution has produced aquatic forms that breathe primarily with lungs and terrestrial forms that lack them completely. The amphiumas of the salamander family Amphiumidae have evolved a completely aquatic life history with a greatly reduced metamorphosis. Amphiumas nonetheless lose their gills before adulthood and then breathe primarily by lungs. They periodically point their nostrils above the surface of the water to get air.
Amphiumas provide a curious contrast
to many species of the family
Plethodontidae that are entirely terrestrial
but have eliminated lungs entirely.
This large family contains more than
220 species, including many familiar
North American salamanders (see Figures
27-6, 27-7, and 27-9). The efficiency
of cutaneous respiration is
increased by penetration of a capillary
network into the epidermis or by thinning
of the epidermis over superficial
dermal capillaries. Cutaneous respiration
is supplemented by pumping air in
and out of the mouth where respiratory
gases are exchanged across the vascularized
membranes of the buccal
(mouth) cavity (buccopharyngeal
breathing). Lungless plethodontids probably originated in swift streams
where lungs would have been a disadvantage
by providing too much buoyancy,
and where water is so cool and
well oxygenated that cutaneous respiration
alone was sufficient for life.
Some plethodontids have aquatic larvae
whose gills are lost at metamorphosis.
Others retain a permanently larval
form with gills throughout life.
Many others are completely terrestrial
and bear the distinction of being the
only vertebrates to have neither lungs
nor gills at any stage of their life history.
It is odd that the most completely
terrestrial lineage of salamanders
evolved in a group that completely
lacks lungs.
Paedomorphosis
A persistent phylogenetic trend observed
in salamander evolution is for
descendants to retain into adulthood
features that were present only in preadult
stages of their ancestors. Some
characteristics of ancestral adult morphology
are consequently eliminated.
This condition is called paedomorphosis (Gr. “child form”; see Organic Evolution). The most dramatic form of
paedomorphosis occurs in those
species that become sexually mature
while retaining their gills, aquatic life
habit, and other larval characteristics.
These nonmetamorphic species are
said to be perennibranchiate (“permanently
gilled”). Mud puppies of the genus Necturus (Figure 27-10), which
live on bottoms of ponds and lakes,
are an extreme example. These and
many other salamanders are obligately
perennibranchiate; they have
never been observed to metamorphose
under any conditions.
Some other species of salamanders reach sexual maturity with larval morphology but, unlike Necturus, may metamorphose to terrestrial forms under certain environmental conditions. Good examples are found in Ambystoma tigrinum and some related species from Mexico and the United States. The gilled individuals are called axolotls (Figure 27-10). Their typical habitat consists of small ponds that can disappear through evaporation in dry weather. When its pond evaporates, an axolotl metamorphoses to a terrestrial form, losing its gills and developing lungs. It can then travel across land in search of new sources of water, to which it must return to reproduce. Axolotls are forced to metamorphose artificially when they are treated with the thyroid hormone, thyroxine (T4). Thyroid hormones (T3 and T4,) are essential for amphibian metamorphosis. The pituitary gland appears not to become fully active in nonmetamorphosing forms, thereby failing to release the hormone thyrotropin, which is required to stimulate the thyroid gland to produce thyroid hormones.
Paedomorphosis takes many different forms in different groups of salamanders. It may affect the body as a whole or may be restricted to one or a few specific structures. The amphiumas mentioned previously lose their gills and activate their lungs before maturity, but they retain many general features of larval body form. Paedomorphosis is important even in terrestrial plethodontids, which never have an aquatic larval stage. We can see the effects of paedomorphosis, for example, in the shapes of the hands and feet of the tropical plethodontid genus Bolitoglossa (Figure 27-11). The ancestral morphology of Bolitoglossa features well-formed digits that grow out from the pad of the hand or foot during development. Some species have enhanced their ability to climb smooth vegetation, such as banana trees, by halting growth of the digits and retaining throughout life a padlike foot. This padlike foot can produce adhesion and suction to attach the salamander to smooth vertical surfaces, and thereby serves an important adaptive function.
Frogs and Toads: Order Anura (Salientia)
The more than 3450 species of frogs and toads that compose the order Anura (Gr. an, without, + oura, tail) are for most people the most familiar amphibians. Anura is an old group, known from the Jurassic period, 150 million years ago. Frogs and toads occupy a great variety of habitats. Their aquatic mode of reproduction and water-permeable skin prevent them from wandering too far from sources of water, however, and their ectothermy bars them from polar and subarctic habitats. The name of the order, Anura, refers to an obvious group characteristic, the absence of tails in adults. Although all pass through a tailed larval stage during development, only the genus Ascaphus contains a tail-like structure in the adult. Frogs and toads are specialized for jumping, as suggested by the alternative order name, Salientia, which means leaping.
We see in the appearance and life habit of their larvae further distinctions between the Anura and Caudata. Eggs of most frogs hatch into a tadpole (“polliwog”), having a long, finned tail, both internal and external gills, no legs, specialized mouthparts for herbivorous feeding (some tadpoles, and salamander larvae, are carnivorous), and a highly specialized internal anatomy. They look and act entirely different from adult frogs. Metamorphosis of a frog tadpole to an adult frog is thus a striking transformation. The perennibranchiate condition never occurs in frogs and toads as it does in salamanders.
Frogs and toads are divided into 21 families. The best-known frog families in North America are Ranidae, which contains most of our familiar frogs (Figure 27-12A), and Hylidae, tree frogs (Figure 27-12B). True toads, belonging to family Bufonidae, have short legs, stout bodies, and thick skins, usually with prominent warts (Figure 27-13). However, the term “toad” is used rather loosely to refer also to more or less terrestrial members of several other families.
The largest anuran is the West African Conraua goliath, which is more than 30 cm long from tip of nose to anus (Figure 27-14). This giant eats animals as big as rats and ducks. The smallest frogs recorded are Eleutherodactylus iberia and Psyllophryne didactyla, measuring less than 1 cm in length; they are also the smallest known tetrapods. These tiny frogs, which can be covered by a dime, are found respectively in Cuba and in the Brazilian rain forest. The largest American frog is the bullfrog, Rana catesbeiana (see Figure 27- 12A), which reaches a head and body length of 20 cm.
Habitats and Distribution
Probably the most abundant frogs are the approximately 260 species of genus Rana (Gr. frog), found throughout the temperate and tropical regions of the world except in New Zealand, the oceanic islands, and southern South America. They usually are found near water, although some, such as wood frogs, R. sylvatica, spend most of their time on damp forest floors. Wood frogs probably return to pools only for breeding in early spring. The larger bullfrogs, R. catesbeiana, and green frogs, R. clamitans, are nearly always found in or near permanent water or swampy regions. Leopard frogs, R. pipiens, have a wider variety of habitats and, with all of their subspecies and forms, are the most widespread of North American frogs. This is the species most commonly used in biology laboratories and for classical electrophysiological research. It has been found in some form in nearly every state, although sparingly represented along the extreme western part of the Pacific coast. It also extends far into northern Canada and as far south as Panama.
Within the range of any species, frogs are often restricted to certain localities (for instance, to specific streams or pools) and may be absent or scarce in similar habitats elsewhere. Pickerel frogs (R. palustris) are especially noteworthy in this respect because they are known to be abundant only in certain localized regions. Recent studies have shown that many populations of frogs worldwide may be suffering declines in numbers and becoming even more patchy than usual in their distributions. In most declining populations causes of decline are unknown.*
Most larger frogs are solitary in their habits except during breeding season. During breeding periods most of them, especially males, are very noisy. Each male usually takes possession of a particular perch near water, where he may remain for hours or even days, trying to attract a female to that spot. At times frogs are mainly silent, and their presence is not detected until they are disturbed. When they enter the water, they dart swiftly and reach the bottom of the pool, where they kick up a cloud of muddy water. In swimming, they hold the forelimbs near the body and kick backward with their webbed hindlimbs, which propel them forward. When they come to the surface to breathe, only the head and foreparts are exposed and, since they usually take advantage of any protective vegetation, they are difficult to see.
During winter months most frogs in temperate climates hibernate in the soft mud of the bottoms of pools and streams. Their life processes are at a very low ebb during their hibernation period, and such energy as they need is derived from glycogen and fat stored in their bodies during the spring and summer months. More terrestrial frogs, such as tree frogs, hibernate in humus of the forest floor. They are tolerant of low temperatures, and many actually survive freezing all extracellular fluid, representing 35% of the body water. Such frost-tolerant frogs prepare for winter by accumulating glucose and glycerol in body fluids, thereby protecting tissues from the normally damaging effects of ice-crystal formation.
Adult frogs have numerous enemies, such as snakes, aquatic birds, turtles, raccoons, and humans; fish prey on tadpoles, and only a few tadpoles survive to maturity. Although usually defenseless, many frogs and toads in the tropics and subtropics are aggressive, jumping and biting at predators. Some defend themselves by feigning death. Most anurans can inflate their lungs so that they are difficult to swallow. When disturbed along the margin of a pond or brook, a frog often remains quite still; when it thinks it is detected, it jumps, not always into the water where enemies may be lurking, but into grassy cover on the bank. When held in the hand, a frog may cease its struggles for an instant to put its captor off guard and then leap violently, voiding its urine. A frog’s best protection is its ability to leap and use of poison glands. Bullfrogs in captivity do not hesitate to snap at tormentors and are capable of inflicting painful bites.
Integument and Coloration
The skin of a frog is thin and moist, and it is attached loosely to the body only at certain points. Histologically the skin is composed of two layers: an outer stratified epidermis and an inner spongy dermis (Figure 27-16). The outer layer of epidermal cells (which is shed periodically when a frog or toad “molts”) contains deposits of keratin, a tough, fibrous protein that provides protection against abrasion and loss of water from the skin. More terrestrial amphibians such as toads have especially heavy deposits of keratin, although amphibian keratin is soft, unlike the hard keratin that forms scales, claws, feathers, horns, and hair of amniotes.
The inner layer of epidermis gives rise to two types of integumentary glands that grow into the loose dermal tissues below. Small mucous glands secrete a protective mucous waterproofing onto the skin surface, and large serous glands produce a whitish, watery poison that is highly irritating to would-be predators. All amphibians produce a skin poison, but its effectiveness varies from species to species and with different predators. The extremely toxic poison of three species of Phyllobates, a genus of small South American dendrobatid frogs, is used by a western Colombian Indian tribe to poison points of blowgun darts. Most species of the family Dendrobatidae produce toxic skin secretions, some of which are among the most lethal animal secretions known, more poisonous even than venoms of sea snakes or any of the most poisonous arachnids.
Skin color in frogs is produced, as in other amphibians, by special pigment cells, chromatophores, located mainly in the dermis. Amphibian chromatophores, like those of many other vertebrates, are branched cells containing pigment that may be concentrated in a small area or dispersed throughout the branching processes to control skin coloration (Figure 27-17). Most amphibians have three types of chromatophores: uppermost in the dermis are xanthophores, containing yellow, orange, or red pigments; beneath these lie iridophores, containing a silvery, light-reflecting pigment; and lowermost are melanophores, containing black or brown melanin. Iridophores act like tiny mirrors, reflecting light back through the xanthophores to produce the brightly conspicuous colors of many tropical frogs. Surprisingly perhaps, green hues so common in North American frogs are produced not by green pigment but by an interaction of xanthophores containing a yellow pigment and underlying iridophores that, by reflecting and scattering the light (Tyndall scattering), produce a blue color. Blue light is filtered by the overlying yellow pigment and thus appears green. Many frogs can adjust their color to blend with their background and thus camouflage themselves (Figure 27-18).
Skeletal and Muscular Systems
In amphibians, as in other vertebrates, a well-developed endoskeleton of bone and cartilage provides a framework for muscles in movement and protection for viscera and nervous systems. Movement onto land and the necessity of transforming paddlelike fins into tetrapod legs capable of supporting the body’s weight introduced a new set of stress and leverage problems. Metamorphosis is most noticeable in anurans, whose entire musculoskeletal system is specialized for jumping and swimming by simultaneous extensor thrusts of the hindlimbs.
The amphibian vertebral column assumes a new role as a support from which the abdomen is slung and to which limbs are attached. Since amphibians move with limbs instead of swimming with serial contractions of the trunk musculature, the vertebral column has lost much of the original flexibility characteristic of fishes. It has become a rigid frame for transmitting force from the hindlimbs to their body. Anurans are further specialized by an extreme shortening of the body. Typical frogs have only nine trunk vertebrae and a rodlike urostyle, which represents several fused caudal vertebrae (coccyx) (Figure 27-19). The limbless caecilians, which obviously have not shared these specializations for tetrapod locomotion, may have as many as 285 vertebrae.
The frog skull is also vastly altered as compared with its vertebrate ancestors; it is much lighter in weight and more flattened in profile and has fewer bones and less ossification. The front part of the skull, wherein are located the nose, eyes, and brain, is better developed, whereas the back of the skull, which contained the gill apparatus in fishes, is much reduced (see Figure 27-19).
The bones and muscles in limbs are of typical tetrapod pattern, with three main joints in each limb (hip, knee, and ankle; or shoulder, elbow, and wrist). The foot is typically fiverayed (pentadactyl) and the hand is four-rayed with both foot and hand having several joints in each of the digits (see Figure 27-19). It is a repetitive system that can be derived from one resembling the bone structure of lobefins, which are distinctly suggestive of amphibian limbs (see Figure 27-1). It is not difficult to imagine how selective pressures through millions of years remodeled ancestral lobe-fins into limbs.
Muscles of the limbs are presumably homologous to the radial muscles that move the fins of fishes, but the muscular arrangement has become so complex in tetrapod limbs that its exact correspondence with fin musculature is unclear. Despite this complexity, we can recognize two major groups of muscles on any limb: an anterior and ventral group that pulls the limb forward and toward the midline (protraction and adduction), and a second set of posterior and dorsal muscles that draws the limb back and away from the body (retraction and abduction).
Trunk musculature, which in fishes is segmentally organized into powerful muscular bands (myomeres) for locomotion by lateral flexion, has been much modified during amphibian evolution. The dorsal (epaxial) muscles are arranged to support the head and brace the vertebral column. The ventral (hypaxial) muscles of the belly are more developed in amphibians than in fishes, since they must support the viscera in air without the buoying assistance of water.
Respiration and Vocalization
Amphibians use three respiratory surfaces for gas exchange in air: skin (cutaneous breathing), mouth (buccal breathing), and lungs. Frogs and toads are more dependent on lung breathing than are salamanders; nevertheless, skin provides an important supplementary avenue for gas exchange in anurans, especially during hibernation in winter. Even under normal conditions when lung breathing predominates, carbon dioxide is lost primarily across the skin while oxygen is absorbed primarily across the lungs.
Lungs are supplied by pulmonary arteries (derived from the sixth aortic arches) and blood returns directly to the left atrium by the pulmonary veins. Frog lungs are ovoid, elastic sacs with their inner surfaces divided into a network of septa that are subdivided into small terminal air chambers called alveoli. Alveoli of frog lungs are much larger than those of amniote vertebrates, and consequently frog lungs have a smaller relative surface available for gas exchange: the respiratory surface of Rana pipiens is about 20 cm2 per cubic centimeter of air contained, compared with 300 cm2 for humans. The problem in lung evolution was not development of a good internal vascular surface, but rather the problem of moving air. A frog is a positive-pressure breather that fills its lungs by forcing air into them; this system contrasts with the negative-pressure system of amniotes. The sequence and explanation of breathing in a frog are shown in Figure 27-20. One can easily follow this sequence in a living frog at rest: rhythmical throat movements of mouth breathing may continue some time before flank movements indicate that the lungs are being emptied and refilled.
Both male and female frogs have vocal cords, but those of males are 552 PART 3 The Diversity of Animal Life much better developed. They are located in the larynx, or voice box. A frog produces sound by passing air back and forth over the vocal cords between the lungs and a large pair of sacs (vocal pouches) in the floor of the mouth. The latter also serve as effective resonators in males, which use their voices to attract mates. Most species utter characteristic sounds that identify them. Many people are familiar with the springtime calls of spring peepers, which produce highpitched sounds surprisingly strident for such tiny frogs. The bass notes of green frogs are banjolike, those of leopard frogs are long and guttural, and bullfrogs produce resonant “jug-o-rum” calls.
Circulation
As in fishes, circulation in amphibians is a closed system of arteries and veins serving a vast peripheral network of capillaries through which blood is forced by the action of a single pressure pump, the heart. The principal changes in circuitry involve the shift from gill to lung breathing. With the elimination of gills, a major obstacle to blood flow was removed from the arterial circuit. But two new problems arose. The first was to provide a blood circuit to the lungs. As we have seen, this problem was solved by converting the sixth aortic arch into pulmonary arteries to serve the lungs and by developing new pulmonary veins for returning oxygenated blood to the heart. The second and evidently more difficult evolutionary problem was to separate pulmonary circulation from the rest of the body’s circulation, so that oxygenated blood from the lungs would be sent to the body and deoxygenated venous blood returning from the body would be sent to the lungs. Solving this problem required a double circulation consisting of separate pulmonary and systemic circuits. Tetrapods solved the problem by evolving a partition down the center of the heart, creating a double pump, one for each circuit. However, partitioning is incomplete in amphibians and most reptilian taxa. Birds and mammals have the most completely divided hearts containing two atria and two ventricles.
Frog hearts (Figure 27-21) have two separate atria and a single undivided ventricle. Blood from the body (systemic circuit) first enters a large receiving chamber, the sinus venosus, which forces blood into the right atrium. The left atrium receives freshly oxygenated blood from the lungs. Both atria contract almost simultaneously, driving both right and left atrial blood into the single ventricle. Although the ventricle is undivided, the blood remains mostly separated, so that when the ventricle contracts, oxygenated pulmonary blood enters the systemic circuit and deoxygenated systemic blood enters the pulmonary circuit. This separation is aided by the spiral valve, which divides the systemic and pulmonary flows in the conus arteriosus (Figure 27-21).
Feeding and Digestion
Frogs are carnivorous, as are most other adult amphibians, and they feed on insects, spiders, worms, slugs, snails, millipedes, and nearly anything else that moves and is small enough to swallow whole. They snap at moving prey with their protrusible tongue, which is attached to the front of the mouth and is free behind. The highly glandular free end of the tongue produces a sticky secretion that adheres to prey. When teeth are present on the premaxillae, maxillae, and vomers, they are used to prevent escape of prey, not for biting or chewing. The digestive tract is relatively short in adult amphibians, a characteristic of most carnivores, and it produces a variety of enzymes for digesting proteins, carbohydrates, and fats.
Larval stages of anurans (tadpoles) are usually herbivorous, feeding on pond algae and other vegetable matter; they have a relatively long digestive tract because their bulky food must be submitted to time-consuming fermentation before useful products can be absorbed.
Nervous System and Special Senses
Three fundamental parts of the brain— forebrain (telencephalon), concerned with the sense of smell; midbrain (mesencephalon), concerned with vision; and hindbrain (rhombencephalon), concerned with hearing and balance— have undergone dramatic developmental trends as the vertebrates moved onto land and improved their environmental awareness. Cephalization increases with emphasis on information processing by the brain and a corresponding loss of independence of the spinal ganglia, which are capable only of stereotyped reflexive behavior. Nonetheless, a headless frog preserves an amazing degree of purposive and highly coordinated behavior. With only the spinal cord intact, it maintains normal body posture and can accurately raise its leg to wipe an irritant from its skin. It will even use the opposite leg if the closer leg is held.
The forebrain (Figure 27-22) contains the olfactory center, which assumes greatly increased importance for detection of dilute airborne odors on land. The sense of smell is in fact one of the dominant special senses in frogs. The remainder of the forebrain, the cerebrum, is of little importance in amphibians. Instead, complex integrative activities of frogs are located in the midbrain optic lobes. The hindbrain is divided into an anterior cerebellum and a posterior medulla. The cerebellum (see Figure 27-22), which is concerned with equilibrium and movement coordination, is not well developed in amphibians, especially in terrestrial species, which stay close to the ground and are not noted for dexterity of movement. The cerebellum becomes vastly developed in the fast-moving birds and mammals. The medulla is really the enlarged anterior end of the spinal cord through which pass all sensory neurons except those of vision and smell. Here are located centers for auditory reflexes, respiration, swallowing, and vasomotor control.
Evolution of a semiterrestrial life by amphibians has necessitated a reordering of sensory receptor priorities on land. The pressure-sensitive lateral line (acousticolateral) system of fishes remains only in aquatic larvae of amphibians and in a few strictly aquatic adult amphibian species. This system can serve no useful purpose on land, because it was designed to detect and localize objects in water by reflected pressure waves. Instead the task of detecting airborne sounds falls on the ear.
A frog’s ear is by amniote standards a simple structure: a middle ear closed externally by a large tympanic membrane (eardrum) and containing a columella (stapes) that transmits vibrations to the inner ear (Figure 27-23). The latter contains a utricle, from which arise three semicircular canals, and a saccule bearing a diverticulum, the lagena. The lagena is partly covered with a tectorial membrane, which in its fine structure is not unlike that of the much more complex mammalian cochlea. In most frogs this structure is sensitive to low-frequency sound energy not greater than 4000 Hz (cycles per second); in bullfrogs the main frequency response is in the 100 to 200 Hz range, which matches the energy of a male frog’s low-pitched call.
Vision is the dominant special sense in many amphibians (the mostly blind caecilians are obvious exceptions). Several modifications of ancestral aquatic eyes were required to adapt them for use in air. Lachrymal glands and eyelids evolved to keep eyes moist, wiped free of dust, and shielded from injury. Since the cornea is exposed to air, it is an important refractive surface, removing much of the burden from the lens of bending light rays and focusing the image on the retina. As in the fishes, accommodation (adjusting focus for near and distant objects) is accomplished by moving the lens. Unlike eyes of most fishes, amphibian eyes at rest are adjusted for distant objects and the lens is moved forward to focus on nearby objects.
A retina contains both rods and cones, the latter providing frogs with color vision. The iris contains welldeveloped circular and radial muscles and can rapidly expand or contract the aperture (pupil) to adjust to changing illumination. The upper lid of the eye is fixed, but the lower one is folded into a transparent nictitating membrane capable of moving across the eye surface (Figure 27-24). Frogs and toads generally possess good vision, a property of crucial importance to animals that rely on quick escape to avoid their numerous predators and on accurate movements to capture rapidly moving prey.
Other sensory receptors include tactile and chemical receptors in skin, taste buds on the tongue and palate, and a well-developed olfactory epithelium lining the nasal cavity.
Reproduction
Because frogs and toads are ectothermic, they breed, feed, and grow only during warm seasons of the year. One of the first drives after the dormant period is breeding. In spring males call vociferously to attract females. When their eggs are mature, females enter water and are clasped by males in a process called amplexus (Figure 27-25). As a female lays eggs, a male discharges sperm over the eggs to fertilize them. After fertilization, the jelly layers absorb water and swell. Eggs are laid in large masses, usually anchored to vegetation.
A fertilized egg (zygote) begins development almost immediately (Figure 27-26). By repeated division (cleavage) an egg is converted into a hollow ball of cells (blastula). The blastula undergoes gastrulation and then continues to differentiate to form an embryo with a tail bud. At 6 to 9 days, depending on temperature, a tadpole hatches from the protective jelly coats that had surrounded the original fertilized egg.
At hatching, a tadpole has a distinct head and body with a compressed tail. Its mouth is located on the ventral side of the head and is provided with horny jaws for feeding by scraping vegetation from hard objects. Behind the mouth is a ventral adhesive disc for clinging to objects. In front of the mouth are two deep pits, which later develop into nostrils. Swellings found on each side of the head later become external gills. There are three pairs of external gills, which later transform into internal gills and become covered with a flap of skin (operculum) on each side. On the right side the operculum completely fuses with the body wall, but on the left side a small opening, the spiracle (L. spiraculum, air hole) remains. Water flows through the spiracle after entering the mouth and passing the internal gills. Hindlegs appear first during metamorphosis, while forelimbs remain temporarily hidden by folds of the operculum. The tail is resorbed, the intestine becomes much shorter, the mouth undergoes a transformation into the adult condition, lungs develop, and gills are resorbed (Figure 27-26). Leopard frogs usually complete metamorphosis within 3 months, whereas bullfrogs take 2 or 3 years to complete the process.
Migration of frogs and toads is correlated with their breeding habits. Males usually return to a pond or stream before females, which they then attract by their calls. Some salamanders also have a strong homing instinct, returning each year to reproduce in the same pool, to which they are guided by olfactory cues. The initial stimulus for migration in many cases is attributable to a seasonal cycle in the gonads plus hormonal changes that increase the frogs’ sensitivity to changes in temperature and humidity.
Classification of Class Amphibia
Order Gymnophiona (jim'no-fy'o-na) (Gr. gymnos, naked, + ophioneos, of a snake) (Apoda): caecilians. Body elongate; limbs and limb girdle absent; mesodermal scales present in skin of some; tail short or absent; 95 to 285 vertebrae; pantropical, 6 families, 34 genera, approximately 160 species.
Order Caudata (caw-dot'uh) (L. caudatus, having a tail) (Urodela): salamanders. Body with head, trunk, and tail; no scales; usually two pairs of equal limbs; 10 to 60 vertebrae; predominantly holarctic; 9 living families, 62 genera, approximately 360 species.
Order Anura (uh-nur'uh) (Gr. an, without, + oura, tail) (Salientia): frogs, toads. Head and trunk fused; no tail; no scales; two pairs of limbs; large mouth; lungs; 6 to 10 vertebrae including urostyle (coccyx); cosmopolitan, predominantly tropical; 21 living families; 301 genera; approximately 3450 species.
The three living amphibian orders comprise more than 4200 species. Most share general adaptations for life on land, including skeletal strengthening and a shifting of special sense priorities from the ancestral lateral line system to the senses of smell and hearing. The olfactory epithelium and the ear are redesigned to improve sensitivities to airborne odors and sounds, respectively.
Nonetheless, most amphibians meet problems of independent life on land only halfway. In the ancestral life history of amphibians, eggs are aquatic and hatch to produce an aquatic larval form that uses gills for respiration. A metamorphosis follows in which gills are lost and lungs, which are present throughout larval life, are then activated for respiration. Many amphibians retain this general pattern but there are some important exceptions. Some salamanders lack a complete metamorphosis and retain a permanently aquatic, larval morphology throughout life. Others live entirely on land and lack the aquatic larval phase completely. Both of these are evolutionarily derived conditions. Some frogs also have acquired a strictly terrestrial existence by eliminating the aquatic larval stage. Some frogs, salamanders, and caecilians that undergo the complete metamorphic life cycle nonetheless remain in water as adults rather than moving onto land during their metamorphosis.
Even the most terrestrial amphibians remain dependent on very moist if not aquatic environments. Their skin is thin, and it requires moisture for protection against desiccation in air. An intact frog loses water nearly as rapidly as a skinless frog. Amphibians also require moderately cool environments. Being ectothermic, their body temperature is determined by and varies with the environment, greatly restricting where they can live. Cool and wet environments are especially important for reproduction. Eggs are not well protected from desiccation, and they must be shed directly into water or onto moist terrestrial surfaces. Completely terrestrial amphibians may lay eggs under logs or rocks, in the moist forest floor, in flooded tree holes, in pockets on the mother’s back (Figure 27-4), or in folds of the body wall. One species of Australian frog even broods its young in its vocal pouch.
We now highlight special characteristics of the three major groups of amphibians. We will expand coverage of general amphibian features when discussing groups in which particular features have been studied most extensively. For most features, this group will be the frogs.
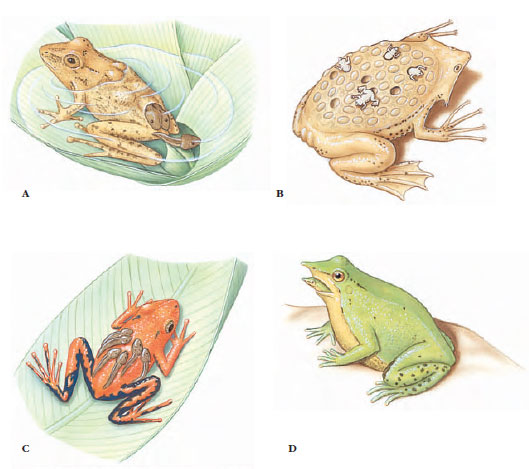 |
| Figure 27-4 Reproductive strategies of anurans. A, Female South American pygmy marsupial frog Flectonotus pygmaeus carries developing larvae in a dorsal pouch. B, Female Surinam frog carries eggs embedded in specialized brooding pouches on the dorsum; froglets emerge and swim away when development is complete. C, Male poison arrow frog Phyllobates bicolor carries tadpoles adhering to its back. D, Tadpoles of a male Darwin’s frog Rhinoderma darwinii develop into froglets in its vocal pouch. When ready to emerge, a froglet crawls into the parent’s mouth, which the parent opens to allow the froglet’s escape. |
Characteristics of Modern Amphibians
- Skeleton mostly bony, with varying numbers of vertebrae; ribs present in some, absent or fused to vertebrae in others; notochord does not persist; exoskeleton absent
- Body forms vary greatly from an elongated trunk with distinct head, neck, and tail to a compact, depressed body with fused head and trunk and no intervening neck
- Limbs usually four (tetrapod), although some are legless; forelimbs of some much smaller than hindlimbs, in others all limbs small and inadequate; webbed feet often present; no true nails or claws; forelimb usually with four digits but sometimes five and sometimes fewer
- Skin smooth and moist with many glands, some of which may be poison glands; pigments cells (chromatophores) common, of considerable variety; no scales, except concealed dermal ones in some
- Mouth usually large with small teeth in upper or both jaws; two nostrils open into anterior part of mouth cavity
- Respiration by lungs (absent in some salamanders), skin, and gills in some, either separately or in combination; external gills in larval forms and may persist throughout life in some
- Circulation with three-chambered heart, two atria and one ventricle, and a double circulation through the heart; skin abundantly supplied with blood vessels
- Ectothermal
- Excretory system of paired mesonephric kidneys; urea main nitrogenous waste
- Ten pairs of cranial nerves
- Separate sexes; fertilization mostly internal in salamanders and caecilians, mostly external in frogs and toads; predominantly oviparous, some ovoviviparous or viviparous; metamorphosis usually present; moderately yolky eggs (mesolecithal) with jellylike membrane coverings
Caecilians: Order Gymnophiona (Apoda)
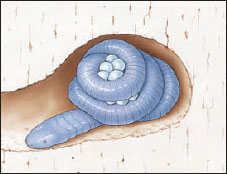 |
| Figure 27-5 Female caecilian coiled around eggs in burrow. |
Salamanders: Order Caudata (Urodela)
As its name suggests, order Caudata (L. caudatus, having a tail) consists of tailed amphibians, approximately 360 species of salamanders. Salamanders are found in almost all northern temperate regions of the world, and they are abundant and diverse in North America. Salamanders are found also in tropical areas of Central and northern South America. Salamanders are typically small; most of the common North American salamanders are less than 15 cm long. Some aquatic forms are considerably longer, and the Japanese giant salamander may exceed 1.5 m in length.
Most salamanders have limbs set at right angles to the body, with forelimbs and hindlimbs of approximately equal size. In some aquatic and burrowing forms, limbs are rudimentary and some may be absent.
Salamanders are carnivorous both as larvae and adults, preying on worms, small arthropods, and small molluscs. Most eat only things that are moving. Since their food is rich in proteins, they do not store great quantities of fat or glycogen. Like all amphibians, they are ectotherms with a low metabolic rate.
 |
| Figure 27-6 Courtship and sperm transfer in pygmy salamanders Desmognathus wrighti. After judging the female’s receptivity by the presence of her chin on his tail base, the male deposits a spermatophore on the ground, then moves forward a few paces. A, The white mass of the sperm atop a gelatinous base is visible at the level of the female’s forelimb. The male moves ahead, the female following until the spermatophore is at the level of her vent. B, The female has recovered the sperm mass in her vent, while the male arches his tail, tilting the female upward and presumably facilitating recovery of the sperm mass. |
Breeding Behavior
Some salamanders are aquatic throughout their life cycle, but most are metamorphic, having aquatic larvae and terrestrial adults that live in moist places under stones and rotten logs. Eggs of most salamanders are fertilized internally, usually after the female recovers in her vent a packet of sperm (spermatophore) that previously has been deposited by the male on a leaf or stick (Figure 27-6). Aquatic species lay their eggs in clusters or stringy masses in water. Their eggs hatch to produce an aquatic larva having external gills and a finlike tail. Completely terrestrial species deposit eggs in small, grapelike clusters under logs or in excavations in soft moist earth, and many species remain to guard the egggs (Figure 27-7). Terrestrial species have direct development: they bypass the larval stage and hatch as miniature versions of their parents. The most complex of salamander life cycles is observed in some American newts,
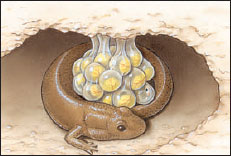 |
| Figure 27-7 Female dusky salamander (Desmognathus sp.) attending eggs. Some salamanders exercise parental care of eggs, which includes rotating eggs and protecting them from fungal infections and predation by various arthropods and other salamanders |
Respiration
Salamanders demonstrate an unusually diverse array of respiratory mechanisms. They share the general amphibian condition of having in their skin extensive vascular nets that serve respiratory exchange of oxygen and carbon dioxide. At various stages of their life history, salamanders also may have external gills, lungs, both, or neither of these. Salamanders with an aquatic larval stage hatch with gills, but lose them later if a metamorphosis occurs. Several diverse lineages of salamanders have evolved permanently aquatic forms that fail to undergo a complete metamorphosis and retain their gills and finlike tail throughout life. Lungs, the most widespread respiratory organ of terrestrial vertebrates, are present from birth in salamanders that have them, and become active following metamorphosis.
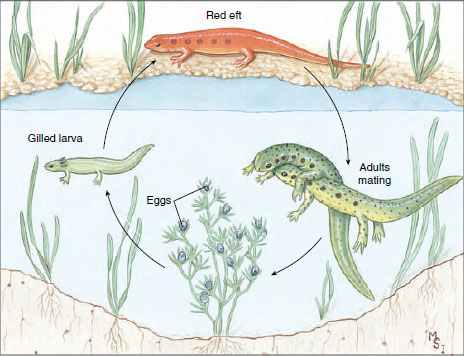 |
| Figure 27-8 Life history of a red-spotted newt, Notophthalmus viridescens of the family Salamandridae. In many habitats the aquatic larva metamorphoses into a brightly colored “red eft” stage, which remains on land from 1 to 3 years before transforming into a secondarily aquatic adult. |
Although we normally associate lungs with terrestrial organisms and gills with aquatic ones, salamander evolution has produced aquatic forms that breathe primarily with lungs and terrestrial forms that lack them completely. The amphiumas of the salamander family Amphiumidae have evolved a completely aquatic life history with a greatly reduced metamorphosis. Amphiumas nonetheless lose their gills before adulthood and then breathe primarily by lungs. They periodically point their nostrils above the surface of the water to get air.
 |
| Figure 27-9 Longtail salamander Eurycea longicauda, a common plethodontid salamander. |
Paedomorphosis
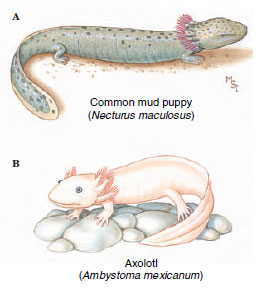 |
| Figure 27-10 Paedomorphosis in salamanders. A, The mud puppy Necturus sp, is a permanently gilled (perennibranchiate) aquatic form. B, An axolotl (Ambystoma tigrinum) may remain permanently gilled, or, should its pond habitat evaporate, metamorphose to a terrestrial form that loses its gills and develops lungs |
Some other species of salamanders reach sexual maturity with larval morphology but, unlike Necturus, may metamorphose to terrestrial forms under certain environmental conditions. Good examples are found in Ambystoma tigrinum and some related species from Mexico and the United States. The gilled individuals are called axolotls (Figure 27-10). Their typical habitat consists of small ponds that can disappear through evaporation in dry weather. When its pond evaporates, an axolotl metamorphoses to a terrestrial form, losing its gills and developing lungs. It can then travel across land in search of new sources of water, to which it must return to reproduce. Axolotls are forced to metamorphose artificially when they are treated with the thyroid hormone, thyroxine (T4). Thyroid hormones (T3 and T4,) are essential for amphibian metamorphosis. The pituitary gland appears not to become fully active in nonmetamorphosing forms, thereby failing to release the hormone thyrotropin, which is required to stimulate the thyroid gland to produce thyroid hormones.
Paedomorphosis takes many different forms in different groups of salamanders. It may affect the body as a whole or may be restricted to one or a few specific structures. The amphiumas mentioned previously lose their gills and activate their lungs before maturity, but they retain many general features of larval body form. Paedomorphosis is important even in terrestrial plethodontids, which never have an aquatic larval stage. We can see the effects of paedomorphosis, for example, in the shapes of the hands and feet of the tropical plethodontid genus Bolitoglossa (Figure 27-11). The ancestral morphology of Bolitoglossa features well-formed digits that grow out from the pad of the hand or foot during development. Some species have enhanced their ability to climb smooth vegetation, such as banana trees, by halting growth of the digits and retaining throughout life a padlike foot. This padlike foot can produce adhesion and suction to attach the salamander to smooth vertical surfaces, and thereby serves an important adaptive function.
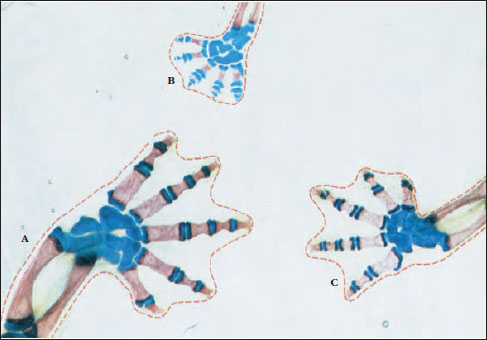 |
| Figure 27-11 Foot structure of representatives of three different species of the tropical plethodontid salamander genus Bolitoglossa. These specimens have been treated chemically to clear the skin and muscles and to stain the bone red and cartilage blue. The species having the most fully ossified and distinct digits (A, C) live primarily on the forest floor. The species having the padlike foot caused by restricted digital growth (B) climbs smooth leaves and stems using the foot surface to produce suction or adhesion for attachment. The padlike foot evolved by paedomorphosis; it was derived evolutionarily by truncating development of the foot to prevent full digital development. |
 |
| Figure 27-12 Two common North American frogs. A, Bullfrog, Rana catesbeiana, largest American frog and mainstay of the frog-leg epicurean market (family Ranidae). B, Green tree frog Hyla cinerea, a common inhabitant of swamps of the southeastern United States (family Hylidae). Note adhesive pads on the feet. |
Frogs and Toads: Order Anura (Salientia)
The more than 3450 species of frogs and toads that compose the order Anura (Gr. an, without, + oura, tail) are for most people the most familiar amphibians. Anura is an old group, known from the Jurassic period, 150 million years ago. Frogs and toads occupy a great variety of habitats. Their aquatic mode of reproduction and water-permeable skin prevent them from wandering too far from sources of water, however, and their ectothermy bars them from polar and subarctic habitats. The name of the order, Anura, refers to an obvious group characteristic, the absence of tails in adults. Although all pass through a tailed larval stage during development, only the genus Ascaphus contains a tail-like structure in the adult. Frogs and toads are specialized for jumping, as suggested by the alternative order name, Salientia, which means leaping.
We see in the appearance and life habit of their larvae further distinctions between the Anura and Caudata. Eggs of most frogs hatch into a tadpole (“polliwog”), having a long, finned tail, both internal and external gills, no legs, specialized mouthparts for herbivorous feeding (some tadpoles, and salamander larvae, are carnivorous), and a highly specialized internal anatomy. They look and act entirely different from adult frogs. Metamorphosis of a frog tadpole to an adult frog is thus a striking transformation. The perennibranchiate condition never occurs in frogs and toads as it does in salamanders.
Frogs and toads are divided into 21 families. The best-known frog families in North America are Ranidae, which contains most of our familiar frogs (Figure 27-12A), and Hylidae, tree frogs (Figure 27-12B). True toads, belonging to family Bufonidae, have short legs, stout bodies, and thick skins, usually with prominent warts (Figure 27-13). However, the term “toad” is used rather loosely to refer also to more or less terrestrial members of several other families.
 |
| Figure 27-13 American toad Bufo americanus (family Bufonidae). This principally nocturnal yet familiar amphibian feeds on large numbers of insect pests and on snails and earthworms. The warty skin contains numerous glands that produce a surprisingly poisonous milky fluid, providing excellent protection from a variety of potential predators. |
The largest anuran is the West African Conraua goliath, which is more than 30 cm long from tip of nose to anus (Figure 27-14). This giant eats animals as big as rats and ducks. The smallest frogs recorded are Eleutherodactylus iberia and Psyllophryne didactyla, measuring less than 1 cm in length; they are also the smallest known tetrapods. These tiny frogs, which can be covered by a dime, are found respectively in Cuba and in the Brazilian rain forest. The largest American frog is the bullfrog, Rana catesbeiana (see Figure 27- 12A), which reaches a head and body length of 20 cm.
 |
| Figure 27-14 Conraua (Gigantorana) goliath (family Ranidae) of West Africa, the world’s largest frog. This specimen weighed 3.3 kg (approximately 71/2 pounds). |
Habitats and Distribution
Probably the most abundant frogs are the approximately 260 species of genus Rana (Gr. frog), found throughout the temperate and tropical regions of the world except in New Zealand, the oceanic islands, and southern South America. They usually are found near water, although some, such as wood frogs, R. sylvatica, spend most of their time on damp forest floors. Wood frogs probably return to pools only for breeding in early spring. The larger bullfrogs, R. catesbeiana, and green frogs, R. clamitans, are nearly always found in or near permanent water or swampy regions. Leopard frogs, R. pipiens, have a wider variety of habitats and, with all of their subspecies and forms, are the most widespread of North American frogs. This is the species most commonly used in biology laboratories and for classical electrophysiological research. It has been found in some form in nearly every state, although sparingly represented along the extreme western part of the Pacific coast. It also extends far into northern Canada and as far south as Panama.
Within the range of any species, frogs are often restricted to certain localities (for instance, to specific streams or pools) and may be absent or scarce in similar habitats elsewhere. Pickerel frogs (R. palustris) are especially noteworthy in this respect because they are known to be abundant only in certain localized regions. Recent studies have shown that many populations of frogs worldwide may be suffering declines in numbers and becoming even more patchy than usual in their distributions. In most declining populations causes of decline are unknown.*
Most larger frogs are solitary in their habits except during breeding season. During breeding periods most of them, especially males, are very noisy. Each male usually takes possession of a particular perch near water, where he may remain for hours or even days, trying to attract a female to that spot. At times frogs are mainly silent, and their presence is not detected until they are disturbed. When they enter the water, they dart swiftly and reach the bottom of the pool, where they kick up a cloud of muddy water. In swimming, they hold the forelimbs near the body and kick backward with their webbed hindlimbs, which propel them forward. When they come to the surface to breathe, only the head and foreparts are exposed and, since they usually take advantage of any protective vegetation, they are difficult to see.
 |
| Figure 27-15 African clawed frog, Xenopus laevis. The claws, an unusual feature in frogs, are on the hind feet. This frog has been introduced into California, where it is considered a serious pest. |
During winter months most frogs in temperate climates hibernate in the soft mud of the bottoms of pools and streams. Their life processes are at a very low ebb during their hibernation period, and such energy as they need is derived from glycogen and fat stored in their bodies during the spring and summer months. More terrestrial frogs, such as tree frogs, hibernate in humus of the forest floor. They are tolerant of low temperatures, and many actually survive freezing all extracellular fluid, representing 35% of the body water. Such frost-tolerant frogs prepare for winter by accumulating glucose and glycerol in body fluids, thereby protecting tissues from the normally damaging effects of ice-crystal formation.
Adult frogs have numerous enemies, such as snakes, aquatic birds, turtles, raccoons, and humans; fish prey on tadpoles, and only a few tadpoles survive to maturity. Although usually defenseless, many frogs and toads in the tropics and subtropics are aggressive, jumping and biting at predators. Some defend themselves by feigning death. Most anurans can inflate their lungs so that they are difficult to swallow. When disturbed along the margin of a pond or brook, a frog often remains quite still; when it thinks it is detected, it jumps, not always into the water where enemies may be lurking, but into grassy cover on the bank. When held in the hand, a frog may cease its struggles for an instant to put its captor off guard and then leap violently, voiding its urine. A frog’s best protection is its ability to leap and use of poison glands. Bullfrogs in captivity do not hesitate to snap at tormentors and are capable of inflicting painful bites.
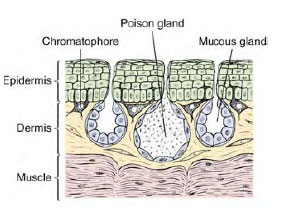 |
| Figure 27-16 Section through frog skin. |
Integument and Coloration
The skin of a frog is thin and moist, and it is attached loosely to the body only at certain points. Histologically the skin is composed of two layers: an outer stratified epidermis and an inner spongy dermis (Figure 27-16). The outer layer of epidermal cells (which is shed periodically when a frog or toad “molts”) contains deposits of keratin, a tough, fibrous protein that provides protection against abrasion and loss of water from the skin. More terrestrial amphibians such as toads have especially heavy deposits of keratin, although amphibian keratin is soft, unlike the hard keratin that forms scales, claws, feathers, horns, and hair of amniotes.
The inner layer of epidermis gives rise to two types of integumentary glands that grow into the loose dermal tissues below. Small mucous glands secrete a protective mucous waterproofing onto the skin surface, and large serous glands produce a whitish, watery poison that is highly irritating to would-be predators. All amphibians produce a skin poison, but its effectiveness varies from species to species and with different predators. The extremely toxic poison of three species of Phyllobates, a genus of small South American dendrobatid frogs, is used by a western Colombian Indian tribe to poison points of blowgun darts. Most species of the family Dendrobatidae produce toxic skin secretions, some of which are among the most lethal animal secretions known, more poisonous even than venoms of sea snakes or any of the most poisonous arachnids.
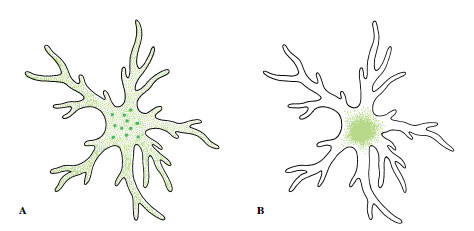
Skin color in frogs is produced, as in other amphibians, by special pigment cells, chromatophores, located mainly in the dermis. Amphibian chromatophores, like those of many other vertebrates, are branched cells containing pigment that may be concentrated in a small area or dispersed throughout the branching processes to control skin coloration (Figure 27-17). Most amphibians have three types of chromatophores: uppermost in the dermis are xanthophores, containing yellow, orange, or red pigments; beneath these lie iridophores, containing a silvery, light-reflecting pigment; and lowermost are melanophores, containing black or brown melanin. Iridophores act like tiny mirrors, reflecting light back through the xanthophores to produce the brightly conspicuous colors of many tropical frogs. Surprisingly perhaps, green hues so common in North American frogs are produced not by green pigment but by an interaction of xanthophores containing a yellow pigment and underlying iridophores that, by reflecting and scattering the light (Tyndall scattering), produce a blue color. Blue light is filtered by the overlying yellow pigment and thus appears green. Many frogs can adjust their color to blend with their background and thus camouflage themselves (Figure 27-18).
 |
 |
| Figure 27-17 Pigment cells (chromatophores). A, Pigment dispersed. B, Pigment concentrated. The pigment cell does not contract or expand; color effects are produced by streaming of cytoplasm, carrying pigment granules into cell branches for maximum color effect or to the center of the cell for minimum effect. Control over dispersal or concentration of pigment is mostly by light stimuli acting through a pituitary hormone. |
Figure 27-18 Cryptic coloration of the gray frog, Hyla versicolor. Camouflage is so good that presence of this frog usually is disclosed only at night by its resonant, flutelike call. |
Skeletal and Muscular Systems
In amphibians, as in other vertebrates, a well-developed endoskeleton of bone and cartilage provides a framework for muscles in movement and protection for viscera and nervous systems. Movement onto land and the necessity of transforming paddlelike fins into tetrapod legs capable of supporting the body’s weight introduced a new set of stress and leverage problems. Metamorphosis is most noticeable in anurans, whose entire musculoskeletal system is specialized for jumping and swimming by simultaneous extensor thrusts of the hindlimbs.
The amphibian vertebral column assumes a new role as a support from which the abdomen is slung and to which limbs are attached. Since amphibians move with limbs instead of swimming with serial contractions of the trunk musculature, the vertebral column has lost much of the original flexibility characteristic of fishes. It has become a rigid frame for transmitting force from the hindlimbs to their body. Anurans are further specialized by an extreme shortening of the body. Typical frogs have only nine trunk vertebrae and a rodlike urostyle, which represents several fused caudal vertebrae (coccyx) (Figure 27-19). The limbless caecilians, which obviously have not shared these specializations for tetrapod locomotion, may have as many as 285 vertebrae.
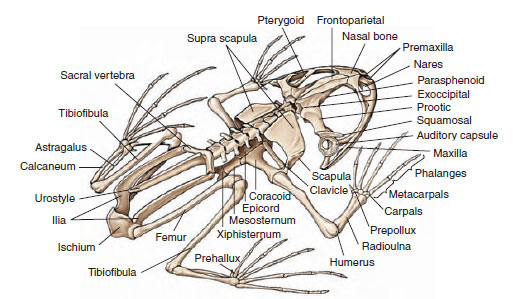 |
| Figure 27-19 Skeleton of a bullfrog, Rana catesbeiana. |
The frog skull is also vastly altered as compared with its vertebrate ancestors; it is much lighter in weight and more flattened in profile and has fewer bones and less ossification. The front part of the skull, wherein are located the nose, eyes, and brain, is better developed, whereas the back of the skull, which contained the gill apparatus in fishes, is much reduced (see Figure 27-19).
The bones and muscles in limbs are of typical tetrapod pattern, with three main joints in each limb (hip, knee, and ankle; or shoulder, elbow, and wrist). The foot is typically fiverayed (pentadactyl) and the hand is four-rayed with both foot and hand having several joints in each of the digits (see Figure 27-19). It is a repetitive system that can be derived from one resembling the bone structure of lobefins, which are distinctly suggestive of amphibian limbs (see Figure 27-1). It is not difficult to imagine how selective pressures through millions of years remodeled ancestral lobe-fins into limbs.
Muscles of the limbs are presumably homologous to the radial muscles that move the fins of fishes, but the muscular arrangement has become so complex in tetrapod limbs that its exact correspondence with fin musculature is unclear. Despite this complexity, we can recognize two major groups of muscles on any limb: an anterior and ventral group that pulls the limb forward and toward the midline (protraction and adduction), and a second set of posterior and dorsal muscles that draws the limb back and away from the body (retraction and abduction).
Trunk musculature, which in fishes is segmentally organized into powerful muscular bands (myomeres) for locomotion by lateral flexion, has been much modified during amphibian evolution. The dorsal (epaxial) muscles are arranged to support the head and brace the vertebral column. The ventral (hypaxial) muscles of the belly are more developed in amphibians than in fishes, since they must support the viscera in air without the buoying assistance of water.
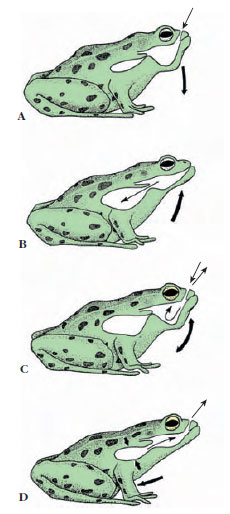 |
| Figure 27-20 Breathing in a frog. Frogs are positive-pressure breathers that fill their lungs by forcing air into them. A, Floor of mouth is lowered, drawing air in through nostrils. B, With nostrils closed and glottis open, the frog forces air into its lungs by elevating floor of mouth. C, Mouth cavity rhythmically ventilates for a period. D, Lungs are emptied by contraction of body-wall musculature and by elastic recoil of lungs. |
Respiration and Vocalization
Amphibians use three respiratory surfaces for gas exchange in air: skin (cutaneous breathing), mouth (buccal breathing), and lungs. Frogs and toads are more dependent on lung breathing than are salamanders; nevertheless, skin provides an important supplementary avenue for gas exchange in anurans, especially during hibernation in winter. Even under normal conditions when lung breathing predominates, carbon dioxide is lost primarily across the skin while oxygen is absorbed primarily across the lungs.
Lungs are supplied by pulmonary arteries (derived from the sixth aortic arches) and blood returns directly to the left atrium by the pulmonary veins. Frog lungs are ovoid, elastic sacs with their inner surfaces divided into a network of septa that are subdivided into small terminal air chambers called alveoli. Alveoli of frog lungs are much larger than those of amniote vertebrates, and consequently frog lungs have a smaller relative surface available for gas exchange: the respiratory surface of Rana pipiens is about 20 cm2 per cubic centimeter of air contained, compared with 300 cm2 for humans. The problem in lung evolution was not development of a good internal vascular surface, but rather the problem of moving air. A frog is a positive-pressure breather that fills its lungs by forcing air into them; this system contrasts with the negative-pressure system of amniotes. The sequence and explanation of breathing in a frog are shown in Figure 27-20. One can easily follow this sequence in a living frog at rest: rhythmical throat movements of mouth breathing may continue some time before flank movements indicate that the lungs are being emptied and refilled.
Both male and female frogs have vocal cords, but those of males are 552 PART 3 The Diversity of Animal Life much better developed. They are located in the larynx, or voice box. A frog produces sound by passing air back and forth over the vocal cords between the lungs and a large pair of sacs (vocal pouches) in the floor of the mouth. The latter also serve as effective resonators in males, which use their voices to attract mates. Most species utter characteristic sounds that identify them. Many people are familiar with the springtime calls of spring peepers, which produce highpitched sounds surprisingly strident for such tiny frogs. The bass notes of green frogs are banjolike, those of leopard frogs are long and guttural, and bullfrogs produce resonant “jug-o-rum” calls.
Circulation
As in fishes, circulation in amphibians is a closed system of arteries and veins serving a vast peripheral network of capillaries through which blood is forced by the action of a single pressure pump, the heart. The principal changes in circuitry involve the shift from gill to lung breathing. With the elimination of gills, a major obstacle to blood flow was removed from the arterial circuit. But two new problems arose. The first was to provide a blood circuit to the lungs. As we have seen, this problem was solved by converting the sixth aortic arch into pulmonary arteries to serve the lungs and by developing new pulmonary veins for returning oxygenated blood to the heart. The second and evidently more difficult evolutionary problem was to separate pulmonary circulation from the rest of the body’s circulation, so that oxygenated blood from the lungs would be sent to the body and deoxygenated venous blood returning from the body would be sent to the lungs. Solving this problem required a double circulation consisting of separate pulmonary and systemic circuits. Tetrapods solved the problem by evolving a partition down the center of the heart, creating a double pump, one for each circuit. However, partitioning is incomplete in amphibians and most reptilian taxa. Birds and mammals have the most completely divided hearts containing two atria and two ventricles.
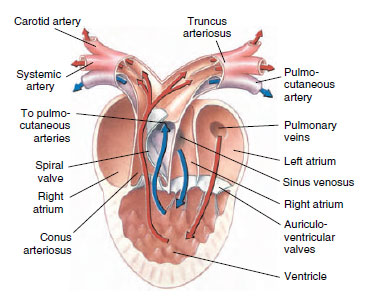 |
| Figure 27-21 Structure of a frog heart. Red arrows, oxygenated blood. Blue arrows, deoxygenated blood. |
Frog hearts (Figure 27-21) have two separate atria and a single undivided ventricle. Blood from the body (systemic circuit) first enters a large receiving chamber, the sinus venosus, which forces blood into the right atrium. The left atrium receives freshly oxygenated blood from the lungs. Both atria contract almost simultaneously, driving both right and left atrial blood into the single ventricle. Although the ventricle is undivided, the blood remains mostly separated, so that when the ventricle contracts, oxygenated pulmonary blood enters the systemic circuit and deoxygenated systemic blood enters the pulmonary circuit. This separation is aided by the spiral valve, which divides the systemic and pulmonary flows in the conus arteriosus (Figure 27-21).
Feeding and Digestion
Frogs are carnivorous, as are most other adult amphibians, and they feed on insects, spiders, worms, slugs, snails, millipedes, and nearly anything else that moves and is small enough to swallow whole. They snap at moving prey with their protrusible tongue, which is attached to the front of the mouth and is free behind. The highly glandular free end of the tongue produces a sticky secretion that adheres to prey. When teeth are present on the premaxillae, maxillae, and vomers, they are used to prevent escape of prey, not for biting or chewing. The digestive tract is relatively short in adult amphibians, a characteristic of most carnivores, and it produces a variety of enzymes for digesting proteins, carbohydrates, and fats.
Larval stages of anurans (tadpoles) are usually herbivorous, feeding on pond algae and other vegetable matter; they have a relatively long digestive tract because their bulky food must be submitted to time-consuming fermentation before useful products can be absorbed.
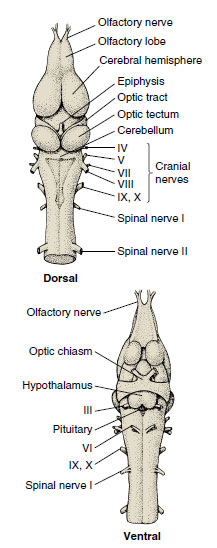 |
| Figure 27-22 Brain of a frog, dorsal and ventral views. |
Nervous System and Special Senses
Three fundamental parts of the brain— forebrain (telencephalon), concerned with the sense of smell; midbrain (mesencephalon), concerned with vision; and hindbrain (rhombencephalon), concerned with hearing and balance— have undergone dramatic developmental trends as the vertebrates moved onto land and improved their environmental awareness. Cephalization increases with emphasis on information processing by the brain and a corresponding loss of independence of the spinal ganglia, which are capable only of stereotyped reflexive behavior. Nonetheless, a headless frog preserves an amazing degree of purposive and highly coordinated behavior. With only the spinal cord intact, it maintains normal body posture and can accurately raise its leg to wipe an irritant from its skin. It will even use the opposite leg if the closer leg is held.
The forebrain (Figure 27-22) contains the olfactory center, which assumes greatly increased importance for detection of dilute airborne odors on land. The sense of smell is in fact one of the dominant special senses in frogs. The remainder of the forebrain, the cerebrum, is of little importance in amphibians. Instead, complex integrative activities of frogs are located in the midbrain optic lobes. The hindbrain is divided into an anterior cerebellum and a posterior medulla. The cerebellum (see Figure 27-22), which is concerned with equilibrium and movement coordination, is not well developed in amphibians, especially in terrestrial species, which stay close to the ground and are not noted for dexterity of movement. The cerebellum becomes vastly developed in the fast-moving birds and mammals. The medulla is really the enlarged anterior end of the spinal cord through which pass all sensory neurons except those of vision and smell. Here are located centers for auditory reflexes, respiration, swallowing, and vasomotor control.
Evolution of a semiterrestrial life by amphibians has necessitated a reordering of sensory receptor priorities on land. The pressure-sensitive lateral line (acousticolateral) system of fishes remains only in aquatic larvae of amphibians and in a few strictly aquatic adult amphibian species. This system can serve no useful purpose on land, because it was designed to detect and localize objects in water by reflected pressure waves. Instead the task of detecting airborne sounds falls on the ear.
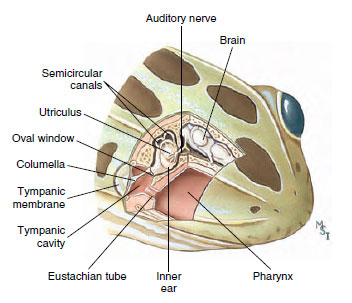 |
| Figure 27-23 Cutaway of frog head showing ear structure. Sound vibrations are transmitted from the tympanic membrane by way of the columella to the inner ear. The eustachian tube allows pressure equilibration between the tympanic cavity and the pharynx. |
A frog’s ear is by amniote standards a simple structure: a middle ear closed externally by a large tympanic membrane (eardrum) and containing a columella (stapes) that transmits vibrations to the inner ear (Figure 27-23). The latter contains a utricle, from which arise three semicircular canals, and a saccule bearing a diverticulum, the lagena. The lagena is partly covered with a tectorial membrane, which in its fine structure is not unlike that of the much more complex mammalian cochlea. In most frogs this structure is sensitive to low-frequency sound energy not greater than 4000 Hz (cycles per second); in bullfrogs the main frequency response is in the 100 to 200 Hz range, which matches the energy of a male frog’s low-pitched call.
Vision is the dominant special sense in many amphibians (the mostly blind caecilians are obvious exceptions). Several modifications of ancestral aquatic eyes were required to adapt them for use in air. Lachrymal glands and eyelids evolved to keep eyes moist, wiped free of dust, and shielded from injury. Since the cornea is exposed to air, it is an important refractive surface, removing much of the burden from the lens of bending light rays and focusing the image on the retina. As in the fishes, accommodation (adjusting focus for near and distant objects) is accomplished by moving the lens. Unlike eyes of most fishes, amphibian eyes at rest are adjusted for distant objects and the lens is moved forward to focus on nearby objects.
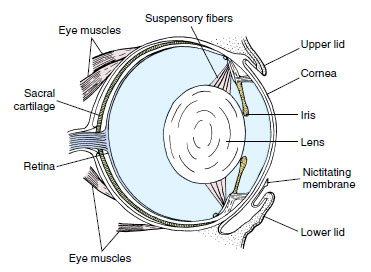 |
| Figure 27-24 Amphibian eye. |
A retina contains both rods and cones, the latter providing frogs with color vision. The iris contains welldeveloped circular and radial muscles and can rapidly expand or contract the aperture (pupil) to adjust to changing illumination. The upper lid of the eye is fixed, but the lower one is folded into a transparent nictitating membrane capable of moving across the eye surface (Figure 27-24). Frogs and toads generally possess good vision, a property of crucial importance to animals that rely on quick escape to avoid their numerous predators and on accurate movements to capture rapidly moving prey.
Other sensory receptors include tactile and chemical receptors in skin, taste buds on the tongue and palate, and a well-developed olfactory epithelium lining the nasal cavity.
Reproduction
Because frogs and toads are ectothermic, they breed, feed, and grow only during warm seasons of the year. One of the first drives after the dormant period is breeding. In spring males call vociferously to attract females. When their eggs are mature, females enter water and are clasped by males in a process called amplexus (Figure 27-25). As a female lays eggs, a male discharges sperm over the eggs to fertilize them. After fertilization, the jelly layers absorb water and swell. Eggs are laid in large masses, usually anchored to vegetation.
 |
| Figure 27-25 A male green frog, Hyla cinerea, clasps a larger female during breeding season in a South Carolina swamp. Clasping (amplexus) is maintained until the female deposits her eggs. Like most tree frogs, these are capable of rapid and marked color changes; the male here, normally green, has darkened during amplexus. |
A fertilized egg (zygote) begins development almost immediately (Figure 27-26). By repeated division (cleavage) an egg is converted into a hollow ball of cells (blastula). The blastula undergoes gastrulation and then continues to differentiate to form an embryo with a tail bud. At 6 to 9 days, depending on temperature, a tadpole hatches from the protective jelly coats that had surrounded the original fertilized egg.
At hatching, a tadpole has a distinct head and body with a compressed tail. Its mouth is located on the ventral side of the head and is provided with horny jaws for feeding by scraping vegetation from hard objects. Behind the mouth is a ventral adhesive disc for clinging to objects. In front of the mouth are two deep pits, which later develop into nostrils. Swellings found on each side of the head later become external gills. There are three pairs of external gills, which later transform into internal gills and become covered with a flap of skin (operculum) on each side. On the right side the operculum completely fuses with the body wall, but on the left side a small opening, the spiracle (L. spiraculum, air hole) remains. Water flows through the spiracle after entering the mouth and passing the internal gills. Hindlegs appear first during metamorphosis, while forelimbs remain temporarily hidden by folds of the operculum. The tail is resorbed, the intestine becomes much shorter, the mouth undergoes a transformation into the adult condition, lungs develop, and gills are resorbed (Figure 27-26). Leopard frogs usually complete metamorphosis within 3 months, whereas bullfrogs take 2 or 3 years to complete the process.
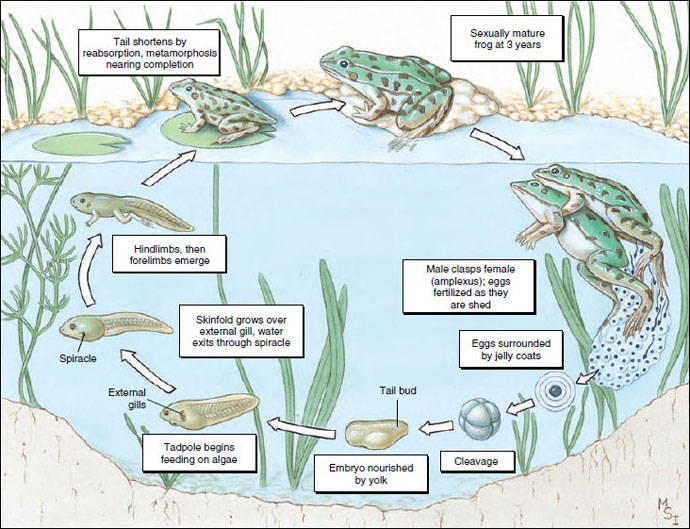 |
| Figure 27-26 Life cycle of a leopard frog. |
Migration of frogs and toads is correlated with their breeding habits. Males usually return to a pond or stream before females, which they then attract by their calls. Some salamanders also have a strong homing instinct, returning each year to reproduce in the same pool, to which they are guided by olfactory cues. The initial stimulus for migration in many cases is attributable to a seasonal cycle in the gonads plus hormonal changes that increase the frogs’ sensitivity to changes in temperature and humidity.
Classification of Class Amphibia
Order Gymnophiona (jim'no-fy'o-na) (Gr. gymnos, naked, + ophioneos, of a snake) (Apoda): caecilians. Body elongate; limbs and limb girdle absent; mesodermal scales present in skin of some; tail short or absent; 95 to 285 vertebrae; pantropical, 6 families, 34 genera, approximately 160 species.
Order Caudata (caw-dot'uh) (L. caudatus, having a tail) (Urodela): salamanders. Body with head, trunk, and tail; no scales; usually two pairs of equal limbs; 10 to 60 vertebrae; predominantly holarctic; 9 living families, 62 genera, approximately 360 species.
Order Anura (uh-nur'uh) (Gr. an, without, + oura, tail) (Salientia): frogs, toads. Head and trunk fused; no tail; no scales; two pairs of limbs; large mouth; lungs; 6 to 10 vertebrae including urostyle (coccyx); cosmopolitan, predominantly tropical; 21 living families; 301 genera; approximately 3450 species.
Support our developers




