Form and Function
Form and Function
The enormous variety, great beauty, and easy availability of shells of molluscs have made shell collecting a popular pastime. However, many amateur shell collectors, even though able to name hundreds of the shells that grace our beaches, know very little about the living animals that created those shells and once lived in them. Reduced to its simplest dimensions, the mollusc body plan may be said to consist of a headfoot portion and a visceral mass portion (Figure 16-2). The head-foot is the more active area, containing the feeding, cephalic sensory, and locomotor organs. It depends primarily on muscular action for its function. The visceral mass is the portion containing digestive, circulatory, respiratory, and reproductive organs, and it depends primarily on ciliary tracts for its functioning. Two folds of skin, outgrowths of the dorsal body wall, make up a protective mantle, or pallium, which encloses a space between the mantle and body wall called the mantle cavity (pallial cavity). The mantle cavity houses gills (ctenidia) or a lung, and in some molluscs the mantle secretes a protective shell over the visceral mass. Modifications of the structures that make up the head-foot and the visceral mass produce the great diversity of patterns observed in Mollusca. Greater emphasis on either the head-foot portion or the visceral mass portion can be observed in various classes of molluscs.
Head-Foot
Most molluscs have well-developed heads, which bear their mouth and some specialized sensory organs. Photosensory receptors range from fairly simple ones to the complex eyes of cephalopods. Tentacles are often present. Within the mouth is a structure unique to molluscs, the radula, and usually posterior to the mouth is the chief locomotor organ, or foot
Radula
The radula is a rasping, protrusible, tonguelike organ found in all molluscs except bivalves and most solenogasters. It is a ribbonlike membrane on which are mounted rows of tiny teeth that point backward (Figure 16-3). Complex muscles move the radula and its supporting cartilages (odontophore) in and out while the membrane is partly rotated over the tips of the cartilages. There may be a few or as many as 250,000 teeth, which, when protruded, can scrape, pierce, tear, or cut. The usual function of the radula is twofold: to rasp off fine particles of food material from hard surfaces and to serve as a conveyor belt for carrying particles in a continuous stream toward the digestive tract. As the radula wears away anteriorly, new rows of teeth are continuously replaced by secretion at its posterior end. The pattern and number of teeth in a row are specific for each species and are used in the classification of molluscs. Very interesting radular specializations, such as for boring through hard materials or for harpooning prey, are found in some forms.
Foot
The molluscan foot (see Figure 16-2) may be variously adapted for locomotion, for attachment to a substratum, or for a combination of functions. It is usually a ventral, solelike structure in which waves of muscular contraction effect a creeping locomotion. However, there are many modifications, such as the attachment disc of limpets, the laterally compressed “hatchet foot” of bivalves, or the siphon for jet propulsion in squids and octopuses. Secreted mucus is often used as an aid to adhesion or as a slime tract by small molluscs that glide on cilia.
In snails and bivalves the foot is extended from the body hydraulically, by engorgement with blood. Burrowing forms can extend the foot into the mud or sand, enlarge it with blood pressure, then use the engorged foot as an anchor to draw the body forward. In pelagic (free-swimming) forms the foot may be modified into winglike parapodia, or thin, mobile fins for swimming.
Visceral Mass
Mantle and Mantle Cavity
The mantle is a sheath of skin, extending
from the visceral mass, that hangs
down on each side of the body, protecting
the soft parts and creating
between itself and the visceral mass a
space called the mantle cavity. The
outer surface of the mantle secretes the
shell.
The mantle cavity (Figure 16-2) plays an enormous role in the life of a mollusc. It usually houses respiratory organs (gills or lung), which develop from the mantle, and the mantle’s own exposed surface serves also for gaseous exchange. Products from the digestive, excretory, and reproductive systems are emptied into the mantle cavity. In aquatic molluscs a continuous current of water, kept moving by surface cilia or by muscular pumping, brings in oxygen and, in some forms, food; flushes out wastes; and carries reproductive products out to the environment. In aquatic forms the mantle is usually equipped with sensory receptors for sampling environmental water. In cephalopods (squids and octopuses) the muscular mantle and its cavity create jet propulsion used in locomotion. Many molluscs can withdraw their head or foot into the mantle cavity, which is surrounded by the shell, for protection.
In primitive form, a mollusc ctenidium (gill) consists of a long, flattened axis extending from the wall of the mantle cavity (Figure 16-4). Many leaflike gill filaments project from the central axis. Water is propelled by cilia between gill filaments, and blood diffuses from an afferent vessel in the central axis through the filament to an efferent vessel. Direction of blood movement is opposite to the direction of water movement, thus establishing a countercurrent exchange mechanism. The two ctenidia are located on opposite sides of the mantle cavity and are arranged so that the cavity is functionally divided into an incurrent chamber and an excurrent chamber. Such gills are found in less derived gastropods, but they are variously modified in many molluscs.
Shell
The shell of a mollusc, when present, is secreted by the mantle and is lined by it. Typically there are three layers (Figure 16-5A). The periostracum is the outer horny layer, composed of an organic substance called conchiolin, which consists of quinone-tanned protein. It helps to protect underlying calcareous layers from erosion by boring organisms. It is secreted by a fold of the mantle edge, and growth occurs only at the margin of the shell. On the older parts of the shell the periostracum often becomes worn away. The middle prismatic layer is composed of densely packed prisms of calcium carbonate laid down in a protein matrix. It is secreted by the glandular margin of the mantle, and increase in shell size occurs at the shell margin as the animal grows. The inner nacreous layer of the shell lies next to the mantle and is secreted continuously by the mantle surface, so that it increases in thickness during the life of the animal. The calcareous nacre is laid down in thin layers. Very thin and wavy layers produce the iridescent mother-of-pearl found in abalones (Haliotis), chambered nautiluses (Nautilus), and many bivalves. Such shells may have 450 to 5000 fine parallel layers of crystalline calcium carbonate (aragonite) for each centimeter of thickness.
Freshwater molluscs usually have a thick periostracum that gives some protection against acids produced in the water by decay of leaf litter. In many marine molluscs the periostracum is relatively thin, and in some it is absent. There is great variation in shell structure. Calcium for the shell comes from environmental water or soil or from food. The first shell appears during the larval period and grows continuously throughout life.
Internal Structure and Function
Gaseous exchange occurs through the body surface, particularly the mantle, and in specialized respiratory organs such as ctenidia, secondary gills, and lungs. There is an open circulatory system with a pumping heart, blood vessels, and blood sinuses. Most cephalopods have a closed blood system with heart, vessels, and capillaries. The digestive tract is complex and highly specialized, according to feeding habits of the various molluscs, and is usually provided with extensive ciliary tracts. Most molluscs have a pair of kidneys (metanephridia, a type of nephridium in which the inner end opens into the coelom by a nephrostome); ducts of the kidneys in many forms also serve for discharge of eggs and sperm.
The nervous system consists of several pairs of ganglia with connecting nerve cords, and it is generally simpler than that of annelids and arthropods. The nervous system contains neurosecretory cells that, at least in certain airbreathing snails, produce a growth hormone and function in osmoregulation. There are various types of highly specialized sense organs.
Reproduction and Life History
Most molluscs are dioecious, although some are hermaphroditic. The freeswimming larva that emerges from the egg in many molluscs is a trochophore, which is also the ancestral larval type of the annelids (Figure 16-6). Direct metamorphosis of the trochophore into a small juvenile, as in chitons, is viewed as ancestral for molluscs, and the intervention of another free-swimming larval stage, a veliger, as in many gastropods and bivalves, is a derived character. The veliger (Figure 16-7) has the beginnings of a foot, shell, and mantle. In many molluscs the trochophore stage occurs in the egg, and a veliger hatches to become the only free-swimming stage. Cephalopods, freshwater and some marine snails, and some freshwater bivalves have no free-swimming larvae, and juveniles hatch directly from eggs.
The enormous variety, great beauty, and easy availability of shells of molluscs have made shell collecting a popular pastime. However, many amateur shell collectors, even though able to name hundreds of the shells that grace our beaches, know very little about the living animals that created those shells and once lived in them. Reduced to its simplest dimensions, the mollusc body plan may be said to consist of a headfoot portion and a visceral mass portion (Figure 16-2). The head-foot is the more active area, containing the feeding, cephalic sensory, and locomotor organs. It depends primarily on muscular action for its function. The visceral mass is the portion containing digestive, circulatory, respiratory, and reproductive organs, and it depends primarily on ciliary tracts for its functioning. Two folds of skin, outgrowths of the dorsal body wall, make up a protective mantle, or pallium, which encloses a space between the mantle and body wall called the mantle cavity (pallial cavity). The mantle cavity houses gills (ctenidia) or a lung, and in some molluscs the mantle secretes a protective shell over the visceral mass. Modifications of the structures that make up the head-foot and the visceral mass produce the great diversity of patterns observed in Mollusca. Greater emphasis on either the head-foot portion or the visceral mass portion can be observed in various classes of molluscs.
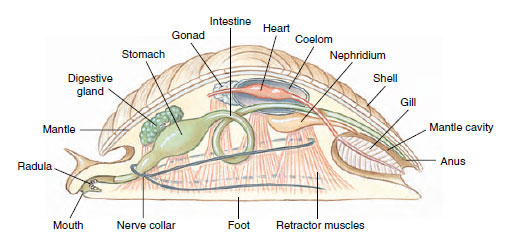 |
| Figure 16-2 Generalized mollusc. Although this construct is often presented as a “hypothetical ancestral mollusc (HAM),” most experts now reject this interpretation. For example, the molluscan ancestor probably was covered with calcareous spicules, rather than a univalve shell. Such a diagram is useful, however, to facilitate description of the general body plan of molluscs. |
Head-Foot
Most molluscs have well-developed heads, which bear their mouth and some specialized sensory organs. Photosensory receptors range from fairly simple ones to the complex eyes of cephalopods. Tentacles are often present. Within the mouth is a structure unique to molluscs, the radula, and usually posterior to the mouth is the chief locomotor organ, or foot
Radula
The radula is a rasping, protrusible, tonguelike organ found in all molluscs except bivalves and most solenogasters. It is a ribbonlike membrane on which are mounted rows of tiny teeth that point backward (Figure 16-3). Complex muscles move the radula and its supporting cartilages (odontophore) in and out while the membrane is partly rotated over the tips of the cartilages. There may be a few or as many as 250,000 teeth, which, when protruded, can scrape, pierce, tear, or cut. The usual function of the radula is twofold: to rasp off fine particles of food material from hard surfaces and to serve as a conveyor belt for carrying particles in a continuous stream toward the digestive tract. As the radula wears away anteriorly, new rows of teeth are continuously replaced by secretion at its posterior end. The pattern and number of teeth in a row are specific for each species and are used in the classification of molluscs. Very interesting radular specializations, such as for boring through hard materials or for harpooning prey, are found in some forms.
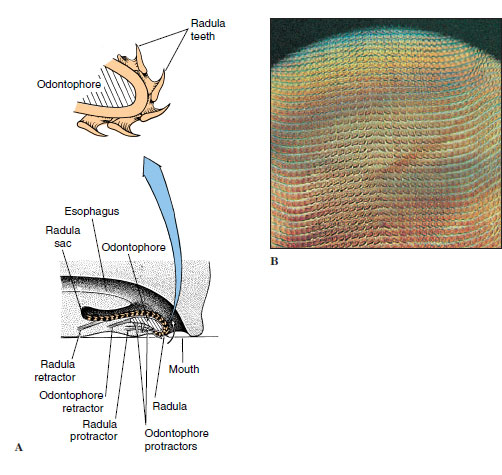 |
| Figure 16-3 A, Diagrammatic longitudinal section of a gastropod head showing a radula and radula sac. The radula moves back and forth over the odontophore cartilage. As the animal grazes, the mouth opens, the odontophore is thrust forward, the radula gives a strong scrape backward bringing food into the pharynx, and the mouth closes. The sequence is repeated rhythmically. As the radula ribbon wears out anteriorly, it is continually replaced posteriorly. B, Radula of a snail prepared for microscopic examination. |
Foot
The molluscan foot (see Figure 16-2) may be variously adapted for locomotion, for attachment to a substratum, or for a combination of functions. It is usually a ventral, solelike structure in which waves of muscular contraction effect a creeping locomotion. However, there are many modifications, such as the attachment disc of limpets, the laterally compressed “hatchet foot” of bivalves, or the siphon for jet propulsion in squids and octopuses. Secreted mucus is often used as an aid to adhesion or as a slime tract by small molluscs that glide on cilia.
In snails and bivalves the foot is extended from the body hydraulically, by engorgement with blood. Burrowing forms can extend the foot into the mud or sand, enlarge it with blood pressure, then use the engorged foot as an anchor to draw the body forward. In pelagic (free-swimming) forms the foot may be modified into winglike parapodia, or thin, mobile fins for swimming.
Visceral Mass
Mantle and Mantle Cavity
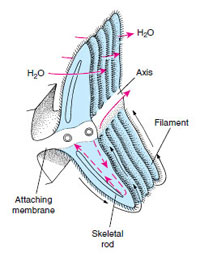 |
| Figure 16-4 Primitive condition of mollusc ctenidium. Circulation of water between gill filaments is by cilia, and blood diffuses through the filament from the afferent vessel to the efferent vessel. Black arrows are ciliary cleansing currents. |
The mantle cavity (Figure 16-2) plays an enormous role in the life of a mollusc. It usually houses respiratory organs (gills or lung), which develop from the mantle, and the mantle’s own exposed surface serves also for gaseous exchange. Products from the digestive, excretory, and reproductive systems are emptied into the mantle cavity. In aquatic molluscs a continuous current of water, kept moving by surface cilia or by muscular pumping, brings in oxygen and, in some forms, food; flushes out wastes; and carries reproductive products out to the environment. In aquatic forms the mantle is usually equipped with sensory receptors for sampling environmental water. In cephalopods (squids and octopuses) the muscular mantle and its cavity create jet propulsion used in locomotion. Many molluscs can withdraw their head or foot into the mantle cavity, which is surrounded by the shell, for protection.
In primitive form, a mollusc ctenidium (gill) consists of a long, flattened axis extending from the wall of the mantle cavity (Figure 16-4). Many leaflike gill filaments project from the central axis. Water is propelled by cilia between gill filaments, and blood diffuses from an afferent vessel in the central axis through the filament to an efferent vessel. Direction of blood movement is opposite to the direction of water movement, thus establishing a countercurrent exchange mechanism. The two ctenidia are located on opposite sides of the mantle cavity and are arranged so that the cavity is functionally divided into an incurrent chamber and an excurrent chamber. Such gills are found in less derived gastropods, but they are variously modified in many molluscs.
Shell
The shell of a mollusc, when present, is secreted by the mantle and is lined by it. Typically there are three layers (Figure 16-5A). The periostracum is the outer horny layer, composed of an organic substance called conchiolin, which consists of quinone-tanned protein. It helps to protect underlying calcareous layers from erosion by boring organisms. It is secreted by a fold of the mantle edge, and growth occurs only at the margin of the shell. On the older parts of the shell the periostracum often becomes worn away. The middle prismatic layer is composed of densely packed prisms of calcium carbonate laid down in a protein matrix. It is secreted by the glandular margin of the mantle, and increase in shell size occurs at the shell margin as the animal grows. The inner nacreous layer of the shell lies next to the mantle and is secreted continuously by the mantle surface, so that it increases in thickness during the life of the animal. The calcareous nacre is laid down in thin layers. Very thin and wavy layers produce the iridescent mother-of-pearl found in abalones (Haliotis), chambered nautiluses (Nautilus), and many bivalves. Such shells may have 450 to 5000 fine parallel layers of crystalline calcium carbonate (aragonite) for each centimeter of thickness.
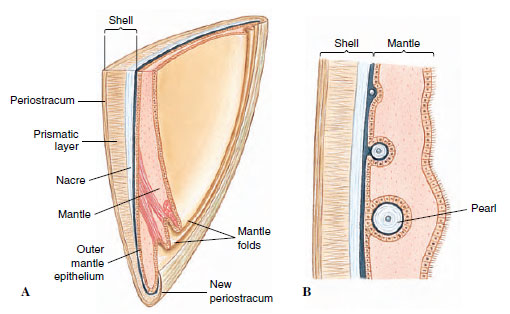 |
| Figure 16-5 A, Diagrammatic vertical section of shell and mantle of a bivalve. The outer mantle epithelium secretes the shell; the inner epithelium is usually ciliated. B, Formation of pearl between mantle and shell as a parasite or bit of sand under the mantle becomes covered with nacre. |
Freshwater molluscs usually have a thick periostracum that gives some protection against acids produced in the water by decay of leaf litter. In many marine molluscs the periostracum is relatively thin, and in some it is absent. There is great variation in shell structure. Calcium for the shell comes from environmental water or soil or from food. The first shell appears during the larval period and grows continuously throughout life.
Internal Structure and Function
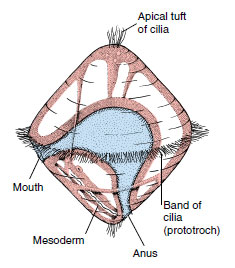 |
| Figure 16-6 Generalized trochophore larva. Molluscs and annelids with primitive embryonic development have trochophore larvae, as do several other phyla. |
Gaseous exchange occurs through the body surface, particularly the mantle, and in specialized respiratory organs such as ctenidia, secondary gills, and lungs. There is an open circulatory system with a pumping heart, blood vessels, and blood sinuses. Most cephalopods have a closed blood system with heart, vessels, and capillaries. The digestive tract is complex and highly specialized, according to feeding habits of the various molluscs, and is usually provided with extensive ciliary tracts. Most molluscs have a pair of kidneys (metanephridia, a type of nephridium in which the inner end opens into the coelom by a nephrostome); ducts of the kidneys in many forms also serve for discharge of eggs and sperm.
The nervous system consists of several pairs of ganglia with connecting nerve cords, and it is generally simpler than that of annelids and arthropods. The nervous system contains neurosecretory cells that, at least in certain airbreathing snails, produce a growth hormone and function in osmoregulation. There are various types of highly specialized sense organs.
Reproduction and Life History
Most molluscs are dioecious, although some are hermaphroditic. The freeswimming larva that emerges from the egg in many molluscs is a trochophore, which is also the ancestral larval type of the annelids (Figure 16-6). Direct metamorphosis of the trochophore into a small juvenile, as in chitons, is viewed as ancestral for molluscs, and the intervention of another free-swimming larval stage, a veliger, as in many gastropods and bivalves, is a derived character. The veliger (Figure 16-7) has the beginnings of a foot, shell, and mantle. In many molluscs the trochophore stage occurs in the egg, and a veliger hatches to become the only free-swimming stage. Cephalopods, freshwater and some marine snails, and some freshwater bivalves have no free-swimming larvae, and juveniles hatch directly from eggs.
Support our developers




