Phylogeny and Adaptive Radiation
Phylogeny and
Adaptive Radiation
The first molluscs probably arose during Precambrian times because fossils attributed to Mollusca have been found in geological strata as old as the early Cambrian period. On the basis of such shared features as spiral cleavage, mesoderm from the 4d blastomere, and trochophore larva, most zoologists have accepted Mollusca as protostomes, allied with the annelids and arthropods. Opinions differ, however, as to whether molluscs were derived from a flatwormlike ancestor independent of annelids, share an ancestor with annelids after the advent of the coelom, or share a metameric common ancestor with annelids. This last hypothesis is strengthened if Neopilina (class Monoplacophora) can be considered metameric, as some scientists have contended. However, it is unlikely that such a successful adaptation as metamerism would have been lost in all later molluscs, and there is no trace of metamerism in development of any known molluscan larva. Therefore most zoologists now suggest that the replication of body parts found in the monoplacophorans is pseudometamerism. The most reasonable hypothesis is that molluscs branched off from the annelid line after the coelom arose but before the advent of metamerism. Some analyses suggest that molluscs and annelids are more closely related to each other than either is to the arthropods. This contention is strengthened by molecular evidence that places annelids and molluscs in Lophotrochozoa and arthropods in Ecdysozoa.
A “hypothetical ancestral mollusc” (see Figure 16-2) was long viewed as representing the original mollusc ancestor, but neither a solid shell nor a broad, crawling foot are now considered universal characters for Mollusca. The primitive ancestral mollusc was probably a more or less wormlike organism with a ventral gliding surface and a dorsal mantle with a chitinous cuticle and calcareous scales (Figure 16-41). It had a posterior mantle cavity with two gills, a radula, a ladderlike nervous system, and an open circulatory system with a heart. Among living molluscs the primitive condition is most nearly approached by caudofoveates, although the foot is reduced to an oral shield in members of this class. Solenogasters have lost the gills, and the foot is represented by the ventral groove. Both these classes probably branched from primitive ancestors before the development of a solid shell, a distinct head with sensory organs, and the ventral muscularized foot. Polyplacophorans probably also branched early from the main lines of molluscan evolution before the veliger was established as the larva. Some workers believe that shells of polyplacophorans are not homologous to shells of other molluscs because they differ structurally and developmentally. Polyplacophora and the remaining classes are sister groups (Figure 16-42).
Some investigators believe that Gastropoda are polyphyletic, perhaps being composed of several groups independently derived from an ancestor shared with monoplacophorans, but cladistic analysis suggests that Gastropoda and Cephalopoda form the sister group to Monoplacophora (see Figure 16-42). Both gastropods and cephalopods have a greatly expanded visceral mass. The mantle cavity was brought toward the head by torsion in gastropods, but in cephalopods the mantle cavity was extended ventrally. Evolution of a chambered shell in cephalopods was a very important contribution to their freedom from the substratum and their ability to swim. Development of their respiratory, circulatory, and nervous systems is correlated with their predatory and swimming habits.
Scaphopods and bivalves have an expanded mantle cavity that essentially envelops the body. Adaptations for burrowing characterize this clade: the spatulate foot and reduction of the head and sense organs.
Most diversity among molluscs is related to their adaptation to different habitats and modes of life and to a wide variety of feeding methods, ranging from sedentary filter feeding to active predation. There are many adaptations for food gathering within the phylum and an enormous variety in radular structure and function, particularly among gastropods.
The versatile glandular mantle has probably shown more plastic adaptive capacity than any other molluscan structure. Besides secreting the shell and forming the mantle cavity, it is variously modified into gills, lungs, siphons, and apertures, and it sometimes functions in locomotion, in the feeding processes, or in a sensory capacity. The shell, too, has undergone a variety of evolutionary adaptations.
Classification of Phylum Mollusca
Useful characteristics for distinction of classes of molluscs are type of foot and type of shell. Several other characteristics are important in particular classes.
The first molluscs probably arose during Precambrian times because fossils attributed to Mollusca have been found in geological strata as old as the early Cambrian period. On the basis of such shared features as spiral cleavage, mesoderm from the 4d blastomere, and trochophore larva, most zoologists have accepted Mollusca as protostomes, allied with the annelids and arthropods. Opinions differ, however, as to whether molluscs were derived from a flatwormlike ancestor independent of annelids, share an ancestor with annelids after the advent of the coelom, or share a metameric common ancestor with annelids. This last hypothesis is strengthened if Neopilina (class Monoplacophora) can be considered metameric, as some scientists have contended. However, it is unlikely that such a successful adaptation as metamerism would have been lost in all later molluscs, and there is no trace of metamerism in development of any known molluscan larva. Therefore most zoologists now suggest that the replication of body parts found in the monoplacophorans is pseudometamerism. The most reasonable hypothesis is that molluscs branched off from the annelid line after the coelom arose but before the advent of metamerism. Some analyses suggest that molluscs and annelids are more closely related to each other than either is to the arthropods. This contention is strengthened by molecular evidence that places annelids and molluscs in Lophotrochozoa and arthropods in Ecdysozoa.
A “hypothetical ancestral mollusc” (see Figure 16-2) was long viewed as representing the original mollusc ancestor, but neither a solid shell nor a broad, crawling foot are now considered universal characters for Mollusca. The primitive ancestral mollusc was probably a more or less wormlike organism with a ventral gliding surface and a dorsal mantle with a chitinous cuticle and calcareous scales (Figure 16-41). It had a posterior mantle cavity with two gills, a radula, a ladderlike nervous system, and an open circulatory system with a heart. Among living molluscs the primitive condition is most nearly approached by caudofoveates, although the foot is reduced to an oral shield in members of this class. Solenogasters have lost the gills, and the foot is represented by the ventral groove. Both these classes probably branched from primitive ancestors before the development of a solid shell, a distinct head with sensory organs, and the ventral muscularized foot. Polyplacophorans probably also branched early from the main lines of molluscan evolution before the veliger was established as the larva. Some workers believe that shells of polyplacophorans are not homologous to shells of other molluscs because they differ structurally and developmentally. Polyplacophora and the remaining classes are sister groups (Figure 16-42).
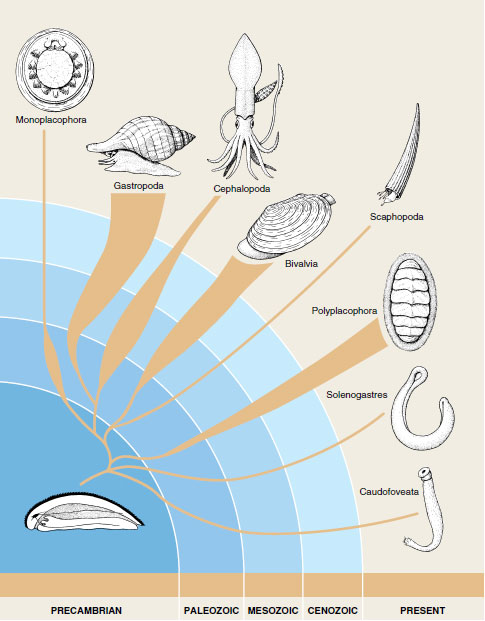 |
| Figure 16-41 Classes of Mollusca, showing their derivations and relative abundance. |
Some investigators believe that Gastropoda are polyphyletic, perhaps being composed of several groups independently derived from an ancestor shared with monoplacophorans, but cladistic analysis suggests that Gastropoda and Cephalopoda form the sister group to Monoplacophora (see Figure 16-42). Both gastropods and cephalopods have a greatly expanded visceral mass. The mantle cavity was brought toward the head by torsion in gastropods, but in cephalopods the mantle cavity was extended ventrally. Evolution of a chambered shell in cephalopods was a very important contribution to their freedom from the substratum and their ability to swim. Development of their respiratory, circulatory, and nervous systems is correlated with their predatory and swimming habits.
Scaphopods and bivalves have an expanded mantle cavity that essentially envelops the body. Adaptations for burrowing characterize this clade: the spatulate foot and reduction of the head and sense organs.
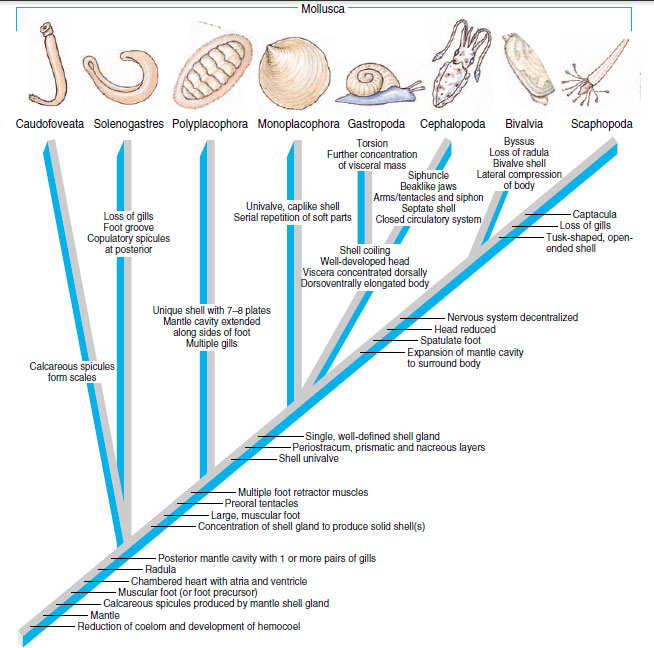 |
| Figure 16-42 Cladogram showing hypothetical relationships among classes of Mollusca. Synapomorphies that identify the various clades are shown, although a number of these have been modified or lost in some descendants. For example, the univalve shell (as well as shell coiling) has been reduced or lost in many gastropods and cephalopods, and many gastropods have undergone detorsion. The bivalve shell of the Bivalvia was derived from an ancestral univalve shell. The byssus is not present in most adult bivalves but functions in larval attachment in many; therefore the byssus is considered a synapomorphy of Bivalvia. |
Most diversity among molluscs is related to their adaptation to different habitats and modes of life and to a wide variety of feeding methods, ranging from sedentary filter feeding to active predation. There are many adaptations for food gathering within the phylum and an enormous variety in radular structure and function, particularly among gastropods.
The versatile glandular mantle has probably shown more plastic adaptive capacity than any other molluscan structure. Besides secreting the shell and forming the mantle cavity, it is variously modified into gills, lungs, siphons, and apertures, and it sometimes functions in locomotion, in the feeding processes, or in a sensory capacity. The shell, too, has undergone a variety of evolutionary adaptations.
Classification of Phylum Mollusca
Useful characteristics for distinction of classes of molluscs are type of foot and type of shell. Several other characteristics are important in particular classes.
-
Class Caudofoveata (kaw´do-fo-ve-at´a) (L. cauda, tail, + fovea, small pit).
Wormlike; shell, head, and excretory
organs absent; radula usually present;
mantle with chitinous cuticle and calcareous
scales; oral pedal shield near
anterior mouth; mantle cavity at posterior
end with pair of gills; sexes
separate; formerly united with
solenogasters in Class Aplacophora.
Examples: Chaetoderma, Limifossor. Class Solenogastres (so-len´o-gas´
trez) (Gr. solen, pipe, + gaster, stomach): solenogasters. Wormlike; shell,
head, and excretory organs absent;
radula present or absent; mantle usually
covered with scales or spicules;
mantle cavity posterior, without true
gills, but sometimes with secondary
respiratory structures; foot represented
by long, narrow, ventral pedal
groove; hermaphroditic. Example: Neomenia.
Class Monoplacophora (mon´o-plakof´o-ra) (Gr. monos, one, + plax, plate, + phora, bearing). Body bilaterally symmetrical with a broad flat foot; a single limpetlike shell; mantle cavity with five or six pairs of gills; large coelomic cavities; radula present; six pairs of nephridia, two of which are gonoducts; separate sexes. Example: Neopilina (see Figure 16-8).
Class Polyplacophora (pol´y-pla-kof´o-ra) (Gr. polys, many, several, + plax, plate, + phora, bearing): chitions. Elongated, dorsoventrally flattened body with reduced head; bilaterally symmetrical; radula present; shell of eight dorsal plates; foot broad and flat; gills multiple along sides of body between foot and mantle edge; sexes usually separate, with a trochophore but no veliger larva. Examples: Mopalia (see Figure 16-10), Tonicella (see Figure 16-1A).
Class Scaphopoda (ska-fop´o-da) (Gr. skaphe, trough, boat, + pous, podos, foot): tusk shells. Body enclosed in a one-piece tubular shell open at both ends; conical foot; mouth with radula and tentacles; head absent; mantle for respiration; sexes separate; trochophore larva. Example: Dentalium (see Figure 16-11).
Class Gastropoda (gas-trop´o-da) (Gr. gaster, stomach, + pous, podos, foot): snails and slugs. Body asymmetrical and shows effects of torsion; body usually in a coiled shell (shell uncoiled or absent in some); head well developed, with radula; foot large and flat; one or two gills, or with mantle modified into secondary gills or a lung; most with single auricle and single nephridium; nervous system with cerebral, pleural, pedal, and visceral ganglia; dioecious or monoecious, some with trochophore, typically with veliger, some without pelagic larva. Examples: Busycon, Polinices (see Figure 16-15B), Physa, Helix, Aplysia (see Figure 16-21). Class Bivalvia (bi-val´ve-a) (L. bi, two, + valva, folding door, valve) (Pelecypoda): bivalves. Body enclosed in a two-lobed mantle; shell of two lateral valves of variable size and form, with dorsal hinge; head greatly reduced, but mouth with labial palps; no radula; no cephalic eyes, a few with eyes on mantle margin; foot usually wedge shaped; gills platelike; sexes usually separate, typically with trochophore and veliger larvae. Examples: Anodonta, Venus, Tagelus (see Figure 16-26), Teredo (see Figure 16-27).
Class Cephalopda (sef ´a-lop´o-da) (Gr. kephale, head, + pous, podos, foot): squids and octopuses. Shell often reduced or absent; head well developed with eyes and a radula; head with arms or tentacles; foot modified into siphon; nervous system of well-developed ganglia, centralized to form a brain; sexes separate, with direct development. Examples: Loligo (see Figure 16-38), Octopus (see Figure 16-1E), Sepia (see Figure 16-37).
Support our developers




