Form and Function
Form and Function
Just as an airplane must be designed and built according to rigid aerodynamic specifications if it is to fly, so too must birds meet stringent structural requirements if they are to stay airborne. All the special adaptations found in flying birds contribute to two things: more power and less weight. Flight by humans became possible when they developed an internal combustion engine and learned how to reduce the weight-topower ratio to a critical point. Birds accomplished flight millions of years ago. But birds must do much more than fly. They must feed themselves and convert food into high-energy fuel; they must escape predators; they must be able to repair their own injuries; they must be able to air-condition themselves when overheated and heat themselves when too cool; and, most important of all, they must reproduce themselves.
Feathers: A feather is very lightweight, yet it possesses remarkable toughness and tensile strength. Most typical of bird feathers are contour feathers, vaned feathers that cover and streamline the bird’s body. A contour feather consists of a hollow quill, or calamus, emerging from a skin follicle, and a shaft, or rachis, which is a continuation of the quill and bears numerous barbs (Figure 29-5). The barbs are arranged in a closely parallel fashion and spread diagonally outward from both sides of the central shaft to form a flat, expansive, webbed surface, the vane. There may be several hundred barbs in the vane.
If a feather is examined with a microscope, each barb appears to be a miniature replica of the feathers with numerous parallel filaments called barbules set in each side of the barb and spreading laterally from it. There may be 600 barbules on each side of a barb, adding up to more than 1 million barbules for the feather. Barbules of one barb overlap barbules of a neighboring barb in a herringbone pattern and are held together with great tenacity by tiny hooks. Should two adjoining barbs become separated—and considerable force is needed to pull the vane apart—they are instantly zipped together again by drawing the feather through the fingertips. The bird, of course, does this preening with its bill, and much of a bird’s time is occupied with preening to keep its feathers in perfect condition.
Types of Feathers
There are different types of bird feathers, serving different functions. Contour feathers (Figure 29-5E) give the bird its outward form and are the type we have already described. Contour feathers that extend beyond the body and are used in flight are called flight feathers. Down feathers (Figure 29-5H) are soft tufts hidden beneath the contour feathers. They are soft because their barbules lack hooks. They are especially abundant on the breast and abdomen of water birds and on young quail and grouse and function principally to conserve heat. Filoplume feathers (Figure 29-5G) are hairlike, degenerate feathers; each is a weak shaft with a tuft of short barbs at the tip. They are the “hairs” of a plucked fowl. They have no known function. Bristles around the mouths of flycatchers and whippoorwills are probably modified filoplumes. A fourth type of highly modified feather, the powder-down feather, is found on herons, bitterns, hawks, and parrots. Tips of these disintegrate as they grow, releasing a talclike powder that helps to waterproof the feathers and give them metallic luster.
Origin and Development
Like the reptiles’ scale to which it is homologous, a feather develops from an epidermal elevation overlying a nourishing dermal core (Figure 29-5A). However, rather than flattening like a scale, the feather bud rolls into a cylinder and sinks into the follicle from which it is growing. During growth, pigments (lipochromes and melanin) are added to the epidermal cells. As the feather enlarges and nears the end of its growth, the soft rachis and barbs are transformed into hard structures by deposition of keratin. The protective sheath splits apart, allowing the end of the feather to protrude and the barbs to unfold.
Molting
When fully grown, a feather, like mammalian hair, is a dead structure. The shedding, or molting, of feathers is a highly orderly process. Except in penguins, which molt all at once, feathers are discarded gradually to avoid the appearance of bare spots. Flight and tail feathers are lost in exact pairs, one from each side, so that balance is maintained (Figure 29-6). Replacements emerge before the next pair is lost, and most birds can continue to fly unimpaired during the molting period; however, many water birds (ducks, geese, loons, and others) lose all their primary feathers at once and are grounded during the molt. Many prepare for molting by moving to isolated bodies of water where they can find food and more easily escape enemies. Nearly all birds molt at least once a year, usually in late summer after the nesting season.
Skeleton
A major structural requirements for flight is a light, yet sturdy, skeleton. As compared with the earliest known bird, Archaeopteryx (Figure 29-7A), the bones of modern birds are phenomenally light, delicate, and laced with air cavities. Such pneumatized bones (Figure 29-8) are nevertheless strong. The skeleton of a frigate bird with a 2.1 m (7-foot) wingspan weighs only 114 grams (4 ounces), less than the weight of all its feathers.
As archosaurs, birds evolved from ancestors with diapsid skulls. However, skulls of modern birds are so specialized that it is difficult to see any trace of the original diapsid condition. The bird skull is built lightly and mostly fused into one piece. The braincase and orbits are large to accommodate a bulging brain and large eyes needed for quick motor coordination and superior vision. Yet, a pigeon skull weighs only 0.21% of its body weight; by comparison the skull of a rat weighs 1.25% of its body weight. As a whole, however, the skeleton of a bird is not lighter than that of a mammal of similar size. The difference is in distribution of mass: whereas the skull and pneumatized wing bones are especially light, the leg bones are heavier than those of mammals. This helps lower the bird’s center of gravity as required for aerodynamic stability.
In Archaeopteryx, both jaws contained teeth set in sockets, an archosaurian characteristic. Modern birds are completely toothless, having instead a horny (keratinous) beak molded around the bony jaws. The mandible is a complex of several bones hinged to provide a double-jointed action that permits the mouth to gape widely. Most birds have kinetic skulls. The attachment of the upper jaw to the skull is flexible; this allows the upper jaw to move slightly, thus increasing the gape. In some birds, parrots for example, the upper jaw is especially flexible because it is hinged to the skull.
The most distinctive feature of the vertebral column is its rigidity. Most vertebrae except the cervicals (neck vertebrae) are fused together, via additional bony structures called uncinate processes (see Figure 29-7), and with the pelvic girdle to form a stiff but light framework to support the legs and provide rigidity for flight. To assist in this rigidity, the ribs are mostly fused with the vertebrae, pectoral girdle, and sternum. Except in flightless birds, the sternum bears a large, thin keel that provides for the attachment of powerful flight muscles. Because Archaeopteryx had no sternum (Figure 29-7B), there was no anchorage for the flight muscles equivalent to that of modern birds. This is one of the principal reasons why Archaeopteryx could not have done any strenuous wingbeating. Archaeopteryx did, however, have a furcula (wishbone) on which enough pectoral muscle could have attached to permit weak flight.
The bones of the forelimbs are highly modified for flight. They are reduced in number, and several are fused together. Despite these alterations, the bird wing is clearly a rearrangement of the basic vertebrate tetrapod limb from which it arose, and all the elements—upper arm, forearm, wrist, and fingers—are represented in modified form (Figure 29-7). The birds’ legs have undergone less pronounced modification than the wings, since they are still designed principally for walking, as well as for perching, scratching, food gathering, and occasionally for swimming, as were those of their archosaurian ancestors.
Muscular System
The locomotor muscles of the wings are relatively massive to meet the demands of flight. Largest of these is the pectoralis, which depresses the wings in flight. Its antagonist is the supracoracoideus muscle, which raises the wing (Figure 29-9). Surprisingly, perhaps, this latter muscle is not located on the backbone (anyone who has been served the back of a chicken knows that it offers little meat) but is positioned under the pectoralis on the breast. It is attached by a tendon to the upper side of the humerus of the wing so that it pulls from below by an ingenious “rope-and-pulley” arrangement. Both pectoralis and supracoracoideus are anchored to the keel. With the main muscle mass low in the body, aerodynamic stability is improved.
The main leg muscle mass is located in the thigh, surrounding the femur, and a smaller mass lies over the tibiotarsus (shank or “drumstick”). Thin but strong tendons extend downward through sleevelike sheaths to the toes. Consequently the feet are nearly devoid of muscles, explaining the thin, delicate appearance of the bird leg. This arrangement places the main muscle mass near the bird’s center of gravity and at the same time allows great agility to the slender, lightweight feet. Because the feet are composed mostly of bone, tendon, and tough, scaly skin, they are highly resistant to damage from freezing. When a bird perches on a branch, an ingenious toe-locking mechanism (Figure 29-10) is activated, which prevents the bird from falling off its perch when asleep. The same mechanism causes the talons of a hawk or owl automatically to sink deeply into its prey as the legs bend under the impact of the strike. The powerful grip of a bird of prey was described by L. Brown
Food, Feeding, and Digestion
In their early evolution, most birds were carnivorous, feeding principally on insects, already well established on the earth’s surface in both variety and numbers long before birds made their appearance. With the advantage of flight, birds could hunt insects on the wing and carry their assault to insect refuges mostly inaccessible to their earthbound tetrapod peers. Today, there is a bird to hunt nearly every insect; they probe the soil, search the bark, scrutinize every leaf and twig, and drill into insect galleries hidden in tree trunks.
Other animal foods (worms, molluscs, crustaceans, fish, frogs, reptiles, mammals, as well as other birds) all found their way into the diet of birds. A very large group, nearly one-fifth of all birds, feeds on nectar. Some birds are omnivores (often termed euryphagous, or “wide-eating” species) that will eat whatever is seasonally abundant. However, omnivorous birds must compete with numerous other omnivores for the same broad spectrum of food. Others are specialists (called stenophagous, or “narroweating” species) that have the pantry to themselves—but at a price. Should the food specialty be reduced or destroyed for some reason (disease, adverse climate, and the like), their very survival may be jeopardized.
The beaks of birds are strongly adapted to specialized food habits— from generalized types such as the strong, pointed beaks of crows, to grotesque, highly specialized ones in flamingoes, pelicans, and avocets (Figure 29-11). The beak of a woodpecker is a straight, hard, chisel-like device. Anchored to a tree trunk with its tail serving as a brace, the woodpecker delivers powerful, rapid blows to excavate nest cavities or expose burrows of wood-boring insects. It then uses its long, flexible, barbed tongue to seek out insects in their galleries. The woodpecker’s skull is especially thick to absorb shock.
How much do birds eat? By a
peculiar twist of reality, the commonplace
“to eat like a bird” is supposed
to signify a diminutive appetite. Yet
birds, because of their intense metabolism,
are voracious feeders. Small
birds with their high metabolic rate
eat more food relative to their body
mass than large birds. This happens
because oxygen consumption increases
only about three-fourths as
rapidly as body weight. For example,
the resting metabolic rate (oxygen consumed per gram of body weight)
of a hummingbird is 12 times that of a
pigeon and 25 times that of a
chicken. A 3 g hummingbird may eat
100% of its body weight in food each
day, an 11 g blue tit about 30%, and a
1880 g domestic chicken, 3.4%. Obviously
the weight of food consumed
also depends on water content of the
food, since water has no nutritive
value. A 57 g Bohemian waxwing was
estimated to eat 170 g of watery Cotoneaster berries in one day—three
times its body weight! Seed-eaters of
equivalent size might eat only 8 g of
dry seeds per day.
Birds process their food rapidly and thoroughly with efficient digestive equipment. A shrike can digest a mouse in 3 hours, and berries will pass completely through the digestive tract of a thrush in just 30 minutes. Because birds lack teeth, foods that require grinding are reduced in the gizzard (see following text). The poorly developed salivary glands mainly secrete mucus for lubricating food and the slender, horn-covered tongue. There are few taste buds, although all birds can taste to some extent. From the short pharynx a relatively long, muscular, elastic esophagus extends to the stomach. Many birds have an enlargement (crop) of the esophagus at its lower end that serves as a storage chamber.
In pigeons, doves, and some parrots, the crop not only stores food but also produces a lipid- and proteinrich “milk,” composed of sloughed epithelial cells of the crop lining. For a few days after hatching, the helpless young are fed regurgitated crop milk by both parents.
The stomach proper consists of two compartments, a proventriculus, which secretes gastric juice, and the muscular gizzard, which is lined with horny plates that serve as millstones for grinding food. To assist in the grinding process, birds swallow coarse, gritty objects or pebbles, which lodge in the gizzard. Certain birds of prey such as owls form pellets of indigestible materials, for example, bones and fur, in the proventriculus by sloughing the gut lining to enclose this material and ejecting it through the mouth. At the junction of the intestine with the rectum are paired ceca; these are well developed in herbivorous birds in which they serve as fermentation chambers. The terminal part of the digestive system is the cloaca, which also receives the genital ducts and ureters.
In young birds the dorsal wall of the cloaca bears the bursa of Fabricius, which processes the B lymphocytes that are important in the immune response.
Circulatory System
The general plan of bird circulation is not greatly different from that of mammals, although their shared derived characteristics were evolved in parallel. The four-chambered heart is large, with strong ventricular walls; thus, birds share with mammals a complete separation of respiratory and systemic circulations. However, the right aortic arch, instead of the left as in the mammals, leads to the dorsal aorta. The two jugular veins in the neck are connected by a cross vein, an adaptation for shunting blood from one jugular to the other as the head rotates. The brachial and pectoral arteries to the wings and breast are unusually large.
The heartbeat is extremely fast, and, as in mammals, there is an inverse relationship between heart rate and body weight. For example, a turkey has a heart rate at rest of about 93 beats per minute, a chicken has a resting rate of 250 beats per minute, and a blackcapped chickadee has 500 beats per minute when asleep, which may increase to a phenomenal 1000 beats per minute during exercise. Blood pressure in birds is roughly equivalent to that in mammals of similar size.
Bird’s blood contains nucleated, biconvex erythrocytes. (Mammals, the only other endothermic vertebrates, have enucleated, biconcave erythrocytes that are somewhat smaller than those of birds.) The phagocytes, or mobile ameboid cells of the blood, are very active and efficient in birds in repairing wounds and destroying microbes.
Respiratory System
The respiratory system of birds differs radically from the lungs of reptiles and mammals and is marvelously adapted for meeting the high metabolic demands of flight. In birds the finest branches of the bronchi, rather than ending in saclike alveoli as in mammals, are developed as tubelike parabronchi through which air flows continuously. Also unique is the extensive system of nine interconnecting air sacs that are located in pairs in the thorax and abdomen and even extend by tiny tubes into the centers of the long bones (Figure 29-12). The air sacs connect to the lungs in such a way that most of the inspired air bypasses the lungs and flows directly into the posterior air sacs, which serve as reservoirs for fresh air. On expiration, this oxygenated air is passed through the lung and collected in the anterior air sacs. From there it flows directly to the outside. Thus, it takes two respiratory cycles for a single breath of air to pass through the respiratory system, allowing for continuous one-way flow through the respiratory exchange chamber, the parabronchi. The airflow sequence is shown in Figure 29-12. The advantage of such a system is obvious: the lungs receive fresh air during both inspiration and expiration. An almost continuous stream of oxygenated air is passed through a system of richly vascularized parabronchi. Although many details of the bird’s respiratory system are not yet understood, it is clearly the most efficient respiratory systems of any vertebrate.
In addition to performing its principal respiratory function, the air sac system helps cool the bird during vigorous exercise. A pigeon, for example, produces about 27 times more heat when flying than when at rest. The air sacs have numerous diverticula that extend inside the larger pneumatic bones of the pectoral and pelvic girdles, wings, and legs. Because they contain warmed air, they provide considerable buoyancy to the bird.
Excretory System
The relatively large paired metanephric kidneys are composed of many thousands of nephrons, each consisting of a renal corpuscle and a nephric tubule. As in other vertebrates, urine is formed by glomerular filtration followed by selective modification of the filtrate in the tubule. Urine passes by way of ureters to the cloaca. There is no urinary bladder.
Birds, like reptiles, excrete their nitrogenous wastes as uric acid, rather than urea, an adaptation that originated with the evolution of the shelled (amniotic) egg. In shelled eggs, all excretory products must remain within the eggshell with the growing embryo. If urea were produced, it would quickly accumulate in solution to toxic levels. Uric acid, however, crystallizes out of solution and can be stored harmlessly within the egg shell. Thus from an embryonic necessity was born an adult virtue. Because of uric acid’s low solubility, a bird can excrete 1 g of uric acid in only 1.5 to 3 ml of water, whereas a mammal may require 60 ml of water to excrete 1 g of urea. The concentration of uric acid occurs almost entirely in the cloaca, where it is combined with fecal material, and the water reabsorbed.
The bird kidney is much less efficient than the mammalian kidney in the removal of solutes, primarily ions of sodium, potassium, and chloride. Most mammals can concentrate solutes to 4 to 8 times that of the blood, and some such as the desert rodents can concentrate urine to nearly 25 times that of the blood. By comparison, most birds concentrate solutes only slightly greater than the blood (the best that any bird can concentrate is about 6 times that of blood).
To compensate for weak soluteconcentrating ability of the kidney, some birds, especially marine birds that must excrete large salt loads from the food they eat and seawater they drink, use extrarenal mechanisms to remove salt from the body. Salt glands, one located above each eye of sea birds (Figure 29-13), can excrete highly concentrated solutions of sodium chloride, up to twice the concentration of seawater. The salt solution runs out the internal or external nostrils, giving gulls, petrels, and other sea birds a perpetual runny nose. The size of the salt gland in some birds depends on how much salt the bird takes in its diet. For example, a race of mallard ducks living a semimarine life in Greenland has salt glands 10 times larger than those of ordinary freshwater mallards.
Nervous and Sensory Systems
The design of a bird’s nervous and sensory system reflects the complex problems of flight and a highly visible existence, in which it must gather food, mate, defend territory, incubate and rear young, and correctly distinguish friend from foe. The brain of a bird has well-developed cerebral hemispheres, cerebellum, and midbrain tectum (optic lobes) (Figure 29-14). The cerebral cortex—chief coordinating center of the mammalian brain—is thin, unfissured, and poorly developed in birds. But the core of the cerebrum, the corpus striatum, has enlarged into the principal integrative center of the brain, controlling such activities as eating, singing, flying, and all complex instinctive reproductive activities. Relatively intelligent birds, such as crows and parrots, have larger cerebral hemispheres than do less intelligent birds such as
chickens and
pigeons. The cerebellum is a crucial
coordinating center where muscleposition
sense, equilibrium sense, and
visual cues are assembled and used to
coordinate movement and balance.
The optic lobes, laterally bulging
structures of the midbrain, form a
visual association apparatus comparable
to the visual cortex of mammals.
Except in flightless birds, ducks, and vultures, the senses of smell and taste are poorly developed in birds. They do, however, have good hearing and superb vision, the keenest in the animal kingdom. As in mammals, the bird ear consists of three regions: (1) external ear, a sound-conducting canal extending to the eardrum, (2) middle ear, containing a rodlike columella that transmits vibrations, and (3) inner ear, where the organ of hearing, the cochlea, is located. The bird cochlea is much shorter than the coiled mammalian cochlea, yet birds can hear roughly the same range of sound frequencies as humans. However, they do not hear high-frequency sounds as well as similar-sized mammals. Actually, the bird ear far surpasses that of humans in capacity to distinguish differences in intensities and to respond to rapid fluctuations in pitch.
The bird eye resembles that of other
vertebrates in gross structure but is relatively
larger; for a given body size, less
spherical, and almost immobile; instead
of turning their eyes, birds turn their
heads with their long and flexible necks
to scan the visual field. The light-sensitive retina (Figure 29-15) is generously
equipped with rods (for dim light
vision) and cones (for good acuity and
color vision). Cones predominate in
diurnal birds, and rods are more numerous
in nocturnal birds. A distinctive feature
of the bird eye is the pecten,
a highly vascularized organ attached
to the retina near the optic nerve and
jutting into the vitreous humor (Figure
29-15). The pecten is thought to
provide nutrients and oxygen to the eye.
It may do more, but its function remains
largely a mystery.
The position of a bird’s eyes in its head is correlated with its life habits. Vegetarians that must avoid predators have eyes placed laterally to give a wide view of the world; predaceous birds such as hawks and owls have eyes directed to the front, allowing more binocular vision for better depth perception. In birds of prey and some others, the fovea, or region of keenest vision on the retina, is placed in a deep pit, which makes it necessary for the bird to focus exactly on the source. Many birds, moreover, have two foveae on the retina (Figure 29-15): the central one for sharp monocular views and the posterior one for binocular vision. Woodcocks can probably see binocularly both forward and backward. The visual acuity of a hawk is about 8 times that of a human (enabling it to see clearly a crouching rabbit more than a mile away), and an owl’s ability to see in dim light is more than 10 times that of a human. Birds have good color vision, especially toward the red end of the spectrum.
Flight
What prompted the evolution of flight in birds, the ability to rise free of earthbound concerns, as almost every human has dreamed of doing? The air was a relatively unexploited habitat stocked with flying insects for food. Flight also offered escape from terrestrial predators and opportunity to travel rapidly and widely to establish new breeding areas and to benefit from year-round favorable climate by migrating north and south with the seasons.
The fossil evidence is too meager to provide us with a recorded history of the origin of bird flight, but it must have happened in one of two ways: birds began to fly by climbing to a high place and gliding down, or by flapping their way into the air from the ground. The “ground-up” hypothesis holds that birds were ground-dwelling runners with primitive wings used to snare insects. With continued enlargement the protowings eventually enabled the running animal to flap its way into the air.
The more widely favored “treesdown” hypothesis suggests that birds passed through an arboreal apprenticeship of tree climbing, leaping through trees, parachuting, gliding, and finally fully powered flight. One thing seems certain: feathers were an absolute requirement for flight. The evolutionary origin of feathers preceded flight; feathers arose for their thermoregulatory role and made possible subsequent evolution of flight. There is absolutely no support for the idea that bird ancestors were originally membrane-winged flyers, like bats, that later developed feathers.
Bird Wing as a Lift Device
To fly, birds must generate lift forces greater than their own mass in order to become airborne and they must provide propulsion in order to move. They use their wings to provide both. In general, the distal part of the wing, the modified hand bones with the attached primaries, acts as a propeller to provide propulsion. Lift is provided by feathers in the more medial part of the wing, the secondaries, associated with the forearm. The wing is streamlined in cross section, with a slightly concave lower surface (cambered) and with small, tight-fitting feathers where the leading edge meets the air (Figure 29-16). Air slips smoothly over the wing, creating lift with minimum drag. Some lift is produced by positive pressure against the undersurface of the wing. But on the upper side, where the airstream must travel farther and faster over the convex surface, a negative pressure is created that provides more than two-thirds of the total lift.
The lift-to-drag ratio of an airfoil is determined by the angle of tilt (angle of attack) and the airspeed (Figure 29-16). A wing carrying a given load can pass through the air at high speed and small angle of attack or at low speed and larger angle of attack. As speed decreases, lift can be increased by increasing the angle of attack, but drag forces also increase. Finally a point is reached (usually around 15 degrees) at which the angle of attack becomes too steep; turbulence appears on the upper surface, lift is destroyed, and stalling occurs. Stalling can be delayed or prevented by placing a wing slot along the leading edge; this structure directs a layer of rapidly moving air across the upper wing surface. Wing slots were and still are used in aircraft traveling at a low speed. In birds, two kinds of wing slots have developed: (1) the alula, or group of small feathers on the thumb (Figures 29-6 and 29-7), which provides a midwing slot, and (2) slotting between the primary feathers, which provides a wing-tip slot. In a number of songbirds, these together provide stall-preventing slots for nearly the entire outer (and aerodynamically more important) half of the wing.
Flapping Flight
Two forces are required for flapping flight: a vertical lifting force to support the bird’s weight, and a horizontal thrusting force to move the bird forward against the resistive forces of friction. Thrust is provided mainly by primary feathers at the wing tips, while secondary feathers of the inner wing, which do not move so far or so fast, act as an airfoil, providing mainly lift. Greatest power is applied on the downstroke. The primary feathers are bent upward and twist to a steep angle of attack, biting into the air like a propeller (Figure 29-17). The entire wing (and the bird’s body) is pulled forward. On the upstroke, the primary feathers bend in the opposite direction so that their upper surfaces twist into a positive angle of attack to produce thrust, just as the lower surfaces did on the downstroke. A powered upstroke is essential for hovering flight, as in hummingbirds (Figure 29-18), and is important for fast, steep takeoffs by small birds with elliptical wings.
Basic Forms of Bird Wings
Bird wings vary in size and form because the successful exploitation of different habitats has imposed special aerodynamic requirements. Four types of bird wings are easily recognized.
Elliptical Wings: Birds that must maneuver in forested habitats, such as sparrows, warblers, doves, woodpeckers, and magpies (Figure 29-19A), have elliptical wings. This type has a low aspect ratio (ratio of length to average width). The wings of the highly maneuverable British Spitfire fighter plane of World War II fame conformed closely to the outline of a sparrows wing. Elliptical wings are slotted between the primary feathers; this arrangement helps prevent stalling during sharp turns, low-speed flight, and frequent landing and takeoff. Each separated primary feather behaves as a narrow wing with a high angle of attack, providing high lift at low speed. The high maneuverability of the elliptical wing is exemplified by the tiny chickadee, which, if frightened, can change course within 0.03 second.
High-Speed Wings: Birds that feed on the wing, such as swallows, hummingbirds, and swifts, or that make long migrations, such as plovers, sandpipers, terns and gulls, (Figure 29-19B), have wings that sweep back and taper to a slender tip. They are rather flat in section, have a moderately high aspect ratio, and lack wing-tip slotting characteristic of elliptical wings. Sweepback and wide separation of the wing tips reduce “tip vortex,” a drag-creating turbulence that tends to develop at wing tips at faster speeds. This type of wing is aerodynamically efficient for high-speed flight but cannot easily keep a bird airborne at low speeds. The fastest birds, such as sandpipers, clocked at 175 km (109 miles) per hour, belong to this group.
Soaring Wings: The oceanic soaring birds have high-aspect ratio wings resembling those of sailplanes. This group includes albatrosses, frigate birds, and gannets (Figure 29-19C). Such long, narrow wings lack wing slots and are adapted for high speed, high lift, and dynamic soaring. They have the highest aerodynamic efficiency of all wings but are less maneuverable than the wide, slotted wings of land soarers. Dynamic soarers exploit the highly reliable sea winds, using adjacent air currents of different velocities.
High-Lift Wings: Vultures, hawks, eagles, owls, and ospreys (Figure 29-19D)—predators that carry heavy loads—have wings with slotting, alulas, and pronounced camber, all of which promote high lift at low speed. Many of these birds are land soarers, with broad, slotted wings that provide the sensitive response and maneuverability required for static soaring in capricious air currents over land.
Just as an airplane must be designed and built according to rigid aerodynamic specifications if it is to fly, so too must birds meet stringent structural requirements if they are to stay airborne. All the special adaptations found in flying birds contribute to two things: more power and less weight. Flight by humans became possible when they developed an internal combustion engine and learned how to reduce the weight-topower ratio to a critical point. Birds accomplished flight millions of years ago. But birds must do much more than fly. They must feed themselves and convert food into high-energy fuel; they must escape predators; they must be able to repair their own injuries; they must be able to air-condition themselves when overheated and heat themselves when too cool; and, most important of all, they must reproduce themselves.
Feathers: A feather is very lightweight, yet it possesses remarkable toughness and tensile strength. Most typical of bird feathers are contour feathers, vaned feathers that cover and streamline the bird’s body. A contour feather consists of a hollow quill, or calamus, emerging from a skin follicle, and a shaft, or rachis, which is a continuation of the quill and bears numerous barbs (Figure 29-5). The barbs are arranged in a closely parallel fashion and spread diagonally outward from both sides of the central shaft to form a flat, expansive, webbed surface, the vane. There may be several hundred barbs in the vane.
If a feather is examined with a microscope, each barb appears to be a miniature replica of the feathers with numerous parallel filaments called barbules set in each side of the barb and spreading laterally from it. There may be 600 barbules on each side of a barb, adding up to more than 1 million barbules for the feather. Barbules of one barb overlap barbules of a neighboring barb in a herringbone pattern and are held together with great tenacity by tiny hooks. Should two adjoining barbs become separated—and considerable force is needed to pull the vane apart—they are instantly zipped together again by drawing the feather through the fingertips. The bird, of course, does this preening with its bill, and much of a bird’s time is occupied with preening to keep its feathers in perfect condition.
Types of Feathers
There are different types of bird feathers, serving different functions. Contour feathers (Figure 29-5E) give the bird its outward form and are the type we have already described. Contour feathers that extend beyond the body and are used in flight are called flight feathers. Down feathers (Figure 29-5H) are soft tufts hidden beneath the contour feathers. They are soft because their barbules lack hooks. They are especially abundant on the breast and abdomen of water birds and on young quail and grouse and function principally to conserve heat. Filoplume feathers (Figure 29-5G) are hairlike, degenerate feathers; each is a weak shaft with a tuft of short barbs at the tip. They are the “hairs” of a plucked fowl. They have no known function. Bristles around the mouths of flycatchers and whippoorwills are probably modified filoplumes. A fourth type of highly modified feather, the powder-down feather, is found on herons, bitterns, hawks, and parrots. Tips of these disintegrate as they grow, releasing a talclike powder that helps to waterproof the feathers and give them metallic luster.
 |
| Figure 29-5 Types of bird feathers and their development. A to E, Successive stages in the development of a vaned, or contour, feather. Growth occurs within a protective sheath, D, that splits open when growth is complete, allowing the mature feather to spread flat. F to H, Other feather varieties, including a pheasant feather with aftershaft, F, filoplumes, G, and down feathers, H. |
Origin and Development
Like the reptiles’ scale to which it is homologous, a feather develops from an epidermal elevation overlying a nourishing dermal core (Figure 29-5A). However, rather than flattening like a scale, the feather bud rolls into a cylinder and sinks into the follicle from which it is growing. During growth, pigments (lipochromes and melanin) are added to the epidermal cells. As the feather enlarges and nears the end of its growth, the soft rachis and barbs are transformed into hard structures by deposition of keratin. The protective sheath splits apart, allowing the end of the feather to protrude and the barbs to unfold.
Molting
When fully grown, a feather, like mammalian hair, is a dead structure. The shedding, or molting, of feathers is a highly orderly process. Except in penguins, which molt all at once, feathers are discarded gradually to avoid the appearance of bare spots. Flight and tail feathers are lost in exact pairs, one from each side, so that balance is maintained (Figure 29-6). Replacements emerge before the next pair is lost, and most birds can continue to fly unimpaired during the molting period; however, many water birds (ducks, geese, loons, and others) lose all their primary feathers at once and are grounded during the molt. Many prepare for molting by moving to isolated bodies of water where they can find food and more easily escape enemies. Nearly all birds molt at least once a year, usually in late summer after the nesting season.
 |
| Figure 29-6 Osprey, Pandion haliaetus (order Falconiformes) lands while holding a freshly-captured fish. Feathers are molted in sequence in exact pairs so that balance is maintained during flight. |
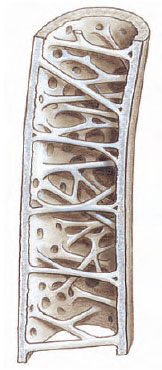 |
| Figure 29-8 Hollow wing bone of a songbird showing the stiffening struts and air spaces that replace bone marrow. Such “pneumatized” bones are remarkably light and strong. |
Skeleton
A major structural requirements for flight is a light, yet sturdy, skeleton. As compared with the earliest known bird, Archaeopteryx (Figure 29-7A), the bones of modern birds are phenomenally light, delicate, and laced with air cavities. Such pneumatized bones (Figure 29-8) are nevertheless strong. The skeleton of a frigate bird with a 2.1 m (7-foot) wingspan weighs only 114 grams (4 ounces), less than the weight of all its feathers.
As archosaurs, birds evolved from ancestors with diapsid skulls. However, skulls of modern birds are so specialized that it is difficult to see any trace of the original diapsid condition. The bird skull is built lightly and mostly fused into one piece. The braincase and orbits are large to accommodate a bulging brain and large eyes needed for quick motor coordination and superior vision. Yet, a pigeon skull weighs only 0.21% of its body weight; by comparison the skull of a rat weighs 1.25% of its body weight. As a whole, however, the skeleton of a bird is not lighter than that of a mammal of similar size. The difference is in distribution of mass: whereas the skull and pneumatized wing bones are especially light, the leg bones are heavier than those of mammals. This helps lower the bird’s center of gravity as required for aerodynamic stability.
In Archaeopteryx, both jaws contained teeth set in sockets, an archosaurian characteristic. Modern birds are completely toothless, having instead a horny (keratinous) beak molded around the bony jaws. The mandible is a complex of several bones hinged to provide a double-jointed action that permits the mouth to gape widely. Most birds have kinetic skulls. The attachment of the upper jaw to the skull is flexible; this allows the upper jaw to move slightly, thus increasing the gape. In some birds, parrots for example, the upper jaw is especially flexible because it is hinged to the skull.
The most distinctive feature of the vertebral column is its rigidity. Most vertebrae except the cervicals (neck vertebrae) are fused together, via additional bony structures called uncinate processes (see Figure 29-7), and with the pelvic girdle to form a stiff but light framework to support the legs and provide rigidity for flight. To assist in this rigidity, the ribs are mostly fused with the vertebrae, pectoral girdle, and sternum. Except in flightless birds, the sternum bears a large, thin keel that provides for the attachment of powerful flight muscles. Because Archaeopteryx had no sternum (Figure 29-7B), there was no anchorage for the flight muscles equivalent to that of modern birds. This is one of the principal reasons why Archaeopteryx could not have done any strenuous wingbeating. Archaeopteryx did, however, have a furcula (wishbone) on which enough pectoral muscle could have attached to permit weak flight.
The bones of the forelimbs are highly modified for flight. They are reduced in number, and several are fused together. Despite these alterations, the bird wing is clearly a rearrangement of the basic vertebrate tetrapod limb from which it arose, and all the elements—upper arm, forearm, wrist, and fingers—are represented in modified form (Figure 29-7). The birds’ legs have undergone less pronounced modification than the wings, since they are still designed principally for walking, as well as for perching, scratching, food gathering, and occasionally for swimming, as were those of their archosaurian ancestors.
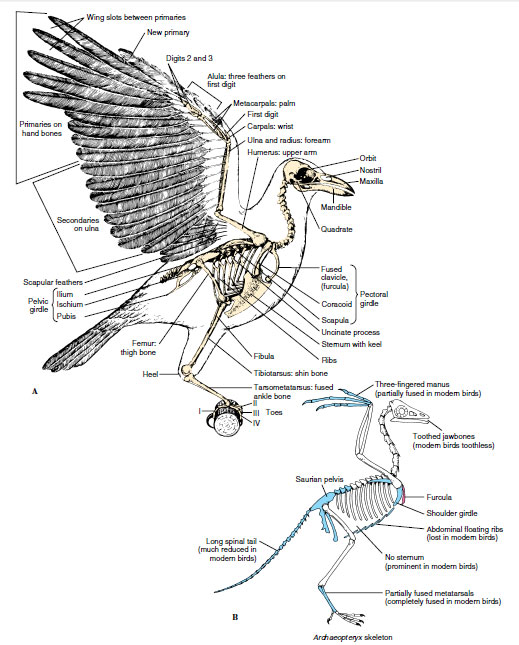 |
| Figure 29-7 A, Skeleton of crow showing portions of the flight feathers. B, Skeleton of Archaeopteryx showing reptilian structures (blue color) that are retained, modified, or lost in modern birds. The furcula (wishbone, red color) was a new avian character. |
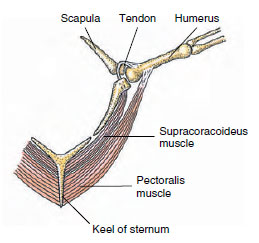 |
| Figure 29-9 Flight muscles of a bird are arranged to keep the center of gravity low in the body. Both major flight muscles are anchored on the sternum keel. Contraction of the pectoralis muscle pulls the wing downward. Then, as the pectoralis relaxes, the supracoracoideus muscle contracts and, acting as a pulley system, pulls the wing upward. |
Muscular System
The locomotor muscles of the wings are relatively massive to meet the demands of flight. Largest of these is the pectoralis, which depresses the wings in flight. Its antagonist is the supracoracoideus muscle, which raises the wing (Figure 29-9). Surprisingly, perhaps, this latter muscle is not located on the backbone (anyone who has been served the back of a chicken knows that it offers little meat) but is positioned under the pectoralis on the breast. It is attached by a tendon to the upper side of the humerus of the wing so that it pulls from below by an ingenious “rope-and-pulley” arrangement. Both pectoralis and supracoracoideus are anchored to the keel. With the main muscle mass low in the body, aerodynamic stability is improved.
The main leg muscle mass is located in the thigh, surrounding the femur, and a smaller mass lies over the tibiotarsus (shank or “drumstick”). Thin but strong tendons extend downward through sleevelike sheaths to the toes. Consequently the feet are nearly devoid of muscles, explaining the thin, delicate appearance of the bird leg. This arrangement places the main muscle mass near the bird’s center of gravity and at the same time allows great agility to the slender, lightweight feet. Because the feet are composed mostly of bone, tendon, and tough, scaly skin, they are highly resistant to damage from freezing. When a bird perches on a branch, an ingenious toe-locking mechanism (Figure 29-10) is activated, which prevents the bird from falling off its perch when asleep. The same mechanism causes the talons of a hawk or owl automatically to sink deeply into its prey as the legs bend under the impact of the strike. The powerful grip of a bird of prey was described by L. Brown
-
When an eagle grips in earnest,
one’s hand becomes numb, and it
is quite impossible to tear it free,
or to loosen the grip of the
eagle’s toes with the other hand.
One just has to wait until the bird
relents, and while waiting one
has ample time to realize that an
animal such as a rabbit would be
quickly paralyzed, unable to draw
breath, and perhaps pierced
through and through by the
talons in such a clutch.
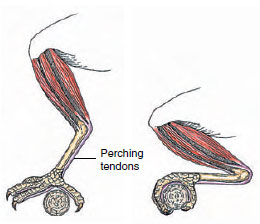 |
| Figure 29-10 Perching mechanism of a bird. When a bird settles on a branch, tendons automatically tighten, closing the toes around the perch. |
Food, Feeding, and Digestion
In their early evolution, most birds were carnivorous, feeding principally on insects, already well established on the earth’s surface in both variety and numbers long before birds made their appearance. With the advantage of flight, birds could hunt insects on the wing and carry their assault to insect refuges mostly inaccessible to their earthbound tetrapod peers. Today, there is a bird to hunt nearly every insect; they probe the soil, search the bark, scrutinize every leaf and twig, and drill into insect galleries hidden in tree trunks.
Other animal foods (worms, molluscs, crustaceans, fish, frogs, reptiles, mammals, as well as other birds) all found their way into the diet of birds. A very large group, nearly one-fifth of all birds, feeds on nectar. Some birds are omnivores (often termed euryphagous, or “wide-eating” species) that will eat whatever is seasonally abundant. However, omnivorous birds must compete with numerous other omnivores for the same broad spectrum of food. Others are specialists (called stenophagous, or “narroweating” species) that have the pantry to themselves—but at a price. Should the food specialty be reduced or destroyed for some reason (disease, adverse climate, and the like), their very survival may be jeopardized.
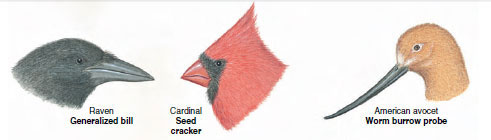
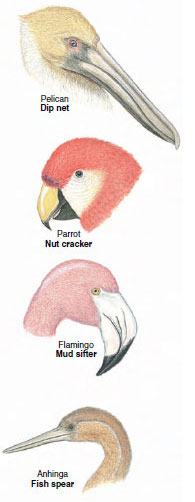
The beaks of birds are strongly adapted to specialized food habits— from generalized types such as the strong, pointed beaks of crows, to grotesque, highly specialized ones in flamingoes, pelicans, and avocets (Figure 29-11). The beak of a woodpecker is a straight, hard, chisel-like device. Anchored to a tree trunk with its tail serving as a brace, the woodpecker delivers powerful, rapid blows to excavate nest cavities or expose burrows of wood-boring insects. It then uses its long, flexible, barbed tongue to seek out insects in their galleries. The woodpecker’s skull is especially thick to absorb shock.
  |
|
| Figure 29-11 Some bills of birds showing variety of adaptations |
|
Birds process their food rapidly and thoroughly with efficient digestive equipment. A shrike can digest a mouse in 3 hours, and berries will pass completely through the digestive tract of a thrush in just 30 minutes. Because birds lack teeth, foods that require grinding are reduced in the gizzard (see following text). The poorly developed salivary glands mainly secrete mucus for lubricating food and the slender, horn-covered tongue. There are few taste buds, although all birds can taste to some extent. From the short pharynx a relatively long, muscular, elastic esophagus extends to the stomach. Many birds have an enlargement (crop) of the esophagus at its lower end that serves as a storage chamber.
In pigeons, doves, and some parrots, the crop not only stores food but also produces a lipid- and proteinrich “milk,” composed of sloughed epithelial cells of the crop lining. For a few days after hatching, the helpless young are fed regurgitated crop milk by both parents.
The stomach proper consists of two compartments, a proventriculus, which secretes gastric juice, and the muscular gizzard, which is lined with horny plates that serve as millstones for grinding food. To assist in the grinding process, birds swallow coarse, gritty objects or pebbles, which lodge in the gizzard. Certain birds of prey such as owls form pellets of indigestible materials, for example, bones and fur, in the proventriculus by sloughing the gut lining to enclose this material and ejecting it through the mouth. At the junction of the intestine with the rectum are paired ceca; these are well developed in herbivorous birds in which they serve as fermentation chambers. The terminal part of the digestive system is the cloaca, which also receives the genital ducts and ureters.
In young birds the dorsal wall of the cloaca bears the bursa of Fabricius, which processes the B lymphocytes that are important in the immune response.
Circulatory System
The general plan of bird circulation is not greatly different from that of mammals, although their shared derived characteristics were evolved in parallel. The four-chambered heart is large, with strong ventricular walls; thus, birds share with mammals a complete separation of respiratory and systemic circulations. However, the right aortic arch, instead of the left as in the mammals, leads to the dorsal aorta. The two jugular veins in the neck are connected by a cross vein, an adaptation for shunting blood from one jugular to the other as the head rotates. The brachial and pectoral arteries to the wings and breast are unusually large.
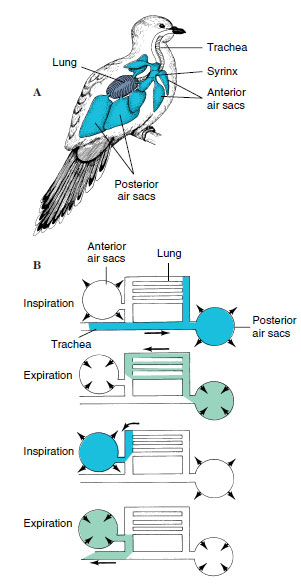 |
| Figure 29-12 Respiratory system of a bird. A, Lungs and air sacs. One side of the bilateral air sac system is shown. B, Movement of a single volume of air through the bird’s respiratory system. Two full respiratory cycles are required to move the air through the system. |
The heartbeat is extremely fast, and, as in mammals, there is an inverse relationship between heart rate and body weight. For example, a turkey has a heart rate at rest of about 93 beats per minute, a chicken has a resting rate of 250 beats per minute, and a blackcapped chickadee has 500 beats per minute when asleep, which may increase to a phenomenal 1000 beats per minute during exercise. Blood pressure in birds is roughly equivalent to that in mammals of similar size.
Bird’s blood contains nucleated, biconvex erythrocytes. (Mammals, the only other endothermic vertebrates, have enucleated, biconcave erythrocytes that are somewhat smaller than those of birds.) The phagocytes, or mobile ameboid cells of the blood, are very active and efficient in birds in repairing wounds and destroying microbes.
Respiratory System
The respiratory system of birds differs radically from the lungs of reptiles and mammals and is marvelously adapted for meeting the high metabolic demands of flight. In birds the finest branches of the bronchi, rather than ending in saclike alveoli as in mammals, are developed as tubelike parabronchi through which air flows continuously. Also unique is the extensive system of nine interconnecting air sacs that are located in pairs in the thorax and abdomen and even extend by tiny tubes into the centers of the long bones (Figure 29-12). The air sacs connect to the lungs in such a way that most of the inspired air bypasses the lungs and flows directly into the posterior air sacs, which serve as reservoirs for fresh air. On expiration, this oxygenated air is passed through the lung and collected in the anterior air sacs. From there it flows directly to the outside. Thus, it takes two respiratory cycles for a single breath of air to pass through the respiratory system, allowing for continuous one-way flow through the respiratory exchange chamber, the parabronchi. The airflow sequence is shown in Figure 29-12. The advantage of such a system is obvious: the lungs receive fresh air during both inspiration and expiration. An almost continuous stream of oxygenated air is passed through a system of richly vascularized parabronchi. Although many details of the bird’s respiratory system are not yet understood, it is clearly the most efficient respiratory systems of any vertebrate.
In addition to performing its principal respiratory function, the air sac system helps cool the bird during vigorous exercise. A pigeon, for example, produces about 27 times more heat when flying than when at rest. The air sacs have numerous diverticula that extend inside the larger pneumatic bones of the pectoral and pelvic girdles, wings, and legs. Because they contain warmed air, they provide considerable buoyancy to the bird.
Excretory System
The relatively large paired metanephric kidneys are composed of many thousands of nephrons, each consisting of a renal corpuscle and a nephric tubule. As in other vertebrates, urine is formed by glomerular filtration followed by selective modification of the filtrate in the tubule. Urine passes by way of ureters to the cloaca. There is no urinary bladder.
Birds, like reptiles, excrete their nitrogenous wastes as uric acid, rather than urea, an adaptation that originated with the evolution of the shelled (amniotic) egg. In shelled eggs, all excretory products must remain within the eggshell with the growing embryo. If urea were produced, it would quickly accumulate in solution to toxic levels. Uric acid, however, crystallizes out of solution and can be stored harmlessly within the egg shell. Thus from an embryonic necessity was born an adult virtue. Because of uric acid’s low solubility, a bird can excrete 1 g of uric acid in only 1.5 to 3 ml of water, whereas a mammal may require 60 ml of water to excrete 1 g of urea. The concentration of uric acid occurs almost entirely in the cloaca, where it is combined with fecal material, and the water reabsorbed.
The bird kidney is much less efficient than the mammalian kidney in the removal of solutes, primarily ions of sodium, potassium, and chloride. Most mammals can concentrate solutes to 4 to 8 times that of the blood, and some such as the desert rodents can concentrate urine to nearly 25 times that of the blood. By comparison, most birds concentrate solutes only slightly greater than the blood (the best that any bird can concentrate is about 6 times that of blood).
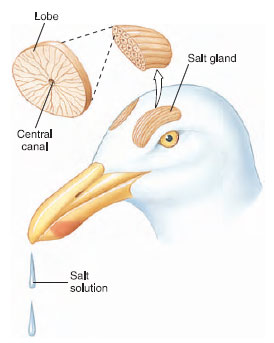 |
| Figure 29-13 Salt glands of a marine bird (gull). One salt gland is located above each eye. Each gland consists of several lobes arranged in parallel. One lobe is shown in cross section, much enlarged. Salt is secreted into many radially arranged tubules, then flows into a central canal that leads into the nose. |
To compensate for weak soluteconcentrating ability of the kidney, some birds, especially marine birds that must excrete large salt loads from the food they eat and seawater they drink, use extrarenal mechanisms to remove salt from the body. Salt glands, one located above each eye of sea birds (Figure 29-13), can excrete highly concentrated solutions of sodium chloride, up to twice the concentration of seawater. The salt solution runs out the internal or external nostrils, giving gulls, petrels, and other sea birds a perpetual runny nose. The size of the salt gland in some birds depends on how much salt the bird takes in its diet. For example, a race of mallard ducks living a semimarine life in Greenland has salt glands 10 times larger than those of ordinary freshwater mallards.
Nervous and Sensory Systems
The design of a bird’s nervous and sensory system reflects the complex problems of flight and a highly visible existence, in which it must gather food, mate, defend territory, incubate and rear young, and correctly distinguish friend from foe. The brain of a bird has well-developed cerebral hemispheres, cerebellum, and midbrain tectum (optic lobes) (Figure 29-14). The cerebral cortex—chief coordinating center of the mammalian brain—is thin, unfissured, and poorly developed in birds. But the core of the cerebrum, the corpus striatum, has enlarged into the principal integrative center of the brain, controlling such activities as eating, singing, flying, and all complex instinctive reproductive activities. Relatively intelligent birds, such as crows and parrots, have larger cerebral hemispheres than do less intelligent birds such as
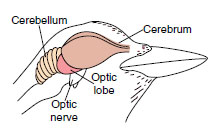 |
| Figure 29-14 Bird brain showing principal divisions. |
Except in flightless birds, ducks, and vultures, the senses of smell and taste are poorly developed in birds. They do, however, have good hearing and superb vision, the keenest in the animal kingdom. As in mammals, the bird ear consists of three regions: (1) external ear, a sound-conducting canal extending to the eardrum, (2) middle ear, containing a rodlike columella that transmits vibrations, and (3) inner ear, where the organ of hearing, the cochlea, is located. The bird cochlea is much shorter than the coiled mammalian cochlea, yet birds can hear roughly the same range of sound frequencies as humans. However, they do not hear high-frequency sounds as well as similar-sized mammals. Actually, the bird ear far surpasses that of humans in capacity to distinguish differences in intensities and to respond to rapid fluctuations in pitch.
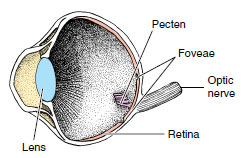 |
| Figure 29-15 |
The position of a bird’s eyes in its head is correlated with its life habits. Vegetarians that must avoid predators have eyes placed laterally to give a wide view of the world; predaceous birds such as hawks and owls have eyes directed to the front, allowing more binocular vision for better depth perception. In birds of prey and some others, the fovea, or region of keenest vision on the retina, is placed in a deep pit, which makes it necessary for the bird to focus exactly on the source. Many birds, moreover, have two foveae on the retina (Figure 29-15): the central one for sharp monocular views and the posterior one for binocular vision. Woodcocks can probably see binocularly both forward and backward. The visual acuity of a hawk is about 8 times that of a human (enabling it to see clearly a crouching rabbit more than a mile away), and an owl’s ability to see in dim light is more than 10 times that of a human. Birds have good color vision, especially toward the red end of the spectrum.
Flight
What prompted the evolution of flight in birds, the ability to rise free of earthbound concerns, as almost every human has dreamed of doing? The air was a relatively unexploited habitat stocked with flying insects for food. Flight also offered escape from terrestrial predators and opportunity to travel rapidly and widely to establish new breeding areas and to benefit from year-round favorable climate by migrating north and south with the seasons.
The fossil evidence is too meager to provide us with a recorded history of the origin of bird flight, but it must have happened in one of two ways: birds began to fly by climbing to a high place and gliding down, or by flapping their way into the air from the ground. The “ground-up” hypothesis holds that birds were ground-dwelling runners with primitive wings used to snare insects. With continued enlargement the protowings eventually enabled the running animal to flap its way into the air.
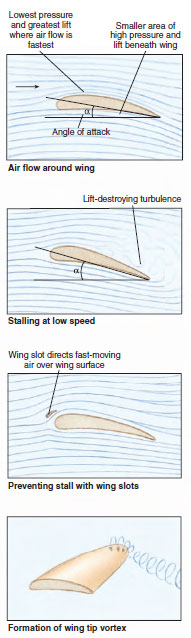 |
| Figure 29-16 Air patterns formed by the airfoil, or wing, moving from right to left. At low speed the angle of attack ( α) must increase to maintain lift but this increases the threat of stalling. The upper figures show how low-speed stalling can be prevented with wing slots. Wing tip vortex (bottom), a turbulence that tends to develop at high speeds, reduces flight efficiency. The effect is reduced in wings that sweep back and taper to a tip. |
The more widely favored “treesdown” hypothesis suggests that birds passed through an arboreal apprenticeship of tree climbing, leaping through trees, parachuting, gliding, and finally fully powered flight. One thing seems certain: feathers were an absolute requirement for flight. The evolutionary origin of feathers preceded flight; feathers arose for their thermoregulatory role and made possible subsequent evolution of flight. There is absolutely no support for the idea that bird ancestors were originally membrane-winged flyers, like bats, that later developed feathers.
Bird Wing as a Lift Device
To fly, birds must generate lift forces greater than their own mass in order to become airborne and they must provide propulsion in order to move. They use their wings to provide both. In general, the distal part of the wing, the modified hand bones with the attached primaries, acts as a propeller to provide propulsion. Lift is provided by feathers in the more medial part of the wing, the secondaries, associated with the forearm. The wing is streamlined in cross section, with a slightly concave lower surface (cambered) and with small, tight-fitting feathers where the leading edge meets the air (Figure 29-16). Air slips smoothly over the wing, creating lift with minimum drag. Some lift is produced by positive pressure against the undersurface of the wing. But on the upper side, where the airstream must travel farther and faster over the convex surface, a negative pressure is created that provides more than two-thirds of the total lift.
The lift-to-drag ratio of an airfoil is determined by the angle of tilt (angle of attack) and the airspeed (Figure 29-16). A wing carrying a given load can pass through the air at high speed and small angle of attack or at low speed and larger angle of attack. As speed decreases, lift can be increased by increasing the angle of attack, but drag forces also increase. Finally a point is reached (usually around 15 degrees) at which the angle of attack becomes too steep; turbulence appears on the upper surface, lift is destroyed, and stalling occurs. Stalling can be delayed or prevented by placing a wing slot along the leading edge; this structure directs a layer of rapidly moving air across the upper wing surface. Wing slots were and still are used in aircraft traveling at a low speed. In birds, two kinds of wing slots have developed: (1) the alula, or group of small feathers on the thumb (Figures 29-6 and 29-7), which provides a midwing slot, and (2) slotting between the primary feathers, which provides a wing-tip slot. In a number of songbirds, these together provide stall-preventing slots for nearly the entire outer (and aerodynamically more important) half of the wing.
Flapping Flight
Two forces are required for flapping flight: a vertical lifting force to support the bird’s weight, and a horizontal thrusting force to move the bird forward against the resistive forces of friction. Thrust is provided mainly by primary feathers at the wing tips, while secondary feathers of the inner wing, which do not move so far or so fast, act as an airfoil, providing mainly lift. Greatest power is applied on the downstroke. The primary feathers are bent upward and twist to a steep angle of attack, biting into the air like a propeller (Figure 29-17). The entire wing (and the bird’s body) is pulled forward. On the upstroke, the primary feathers bend in the opposite direction so that their upper surfaces twist into a positive angle of attack to produce thrust, just as the lower surfaces did on the downstroke. A powered upstroke is essential for hovering flight, as in hummingbirds (Figure 29-18), and is important for fast, steep takeoffs by small birds with elliptical wings.
 |
| Figure 29-17 In normal flapping flight of strong fliers like ducks, the wings sweep downward and forward fully extended. Thrust is provided by the primary feathers at the wing tips. To begin the upbeat, the wing is bent, bringing it upward and backward. The wing then extends, ready for the next downbeat. |
 |
| Figure 29-18 The secret of a hummingbird’s ability to change direction instantly, or hang motionless in the air while sipping nectar from a flower, lies in its wing structure. The wing is nearly rigid, but hinged at the shoulder by a swivel joint and powered by a supracoracoideus muscle that is unusually large for the bird’s size. When hovering the wing moves in a sculling motion. The leading edge of the wing moves forward on the forward stroke, then swivels nearly 180 degrees at the shoulder to move backward on the backstroke. The effect is to provide lift without propulsion on both forward and backstrokes. |
Basic Forms of Bird Wings
Bird wings vary in size and form because the successful exploitation of different habitats has imposed special aerodynamic requirements. Four types of bird wings are easily recognized.
Elliptical Wings: Birds that must maneuver in forested habitats, such as sparrows, warblers, doves, woodpeckers, and magpies (Figure 29-19A), have elliptical wings. This type has a low aspect ratio (ratio of length to average width). The wings of the highly maneuverable British Spitfire fighter plane of World War II fame conformed closely to the outline of a sparrows wing. Elliptical wings are slotted between the primary feathers; this arrangement helps prevent stalling during sharp turns, low-speed flight, and frequent landing and takeoff. Each separated primary feather behaves as a narrow wing with a high angle of attack, providing high lift at low speed. The high maneuverability of the elliptical wing is exemplified by the tiny chickadee, which, if frightened, can change course within 0.03 second.
High-Speed Wings: Birds that feed on the wing, such as swallows, hummingbirds, and swifts, or that make long migrations, such as plovers, sandpipers, terns and gulls, (Figure 29-19B), have wings that sweep back and taper to a slender tip. They are rather flat in section, have a moderately high aspect ratio, and lack wing-tip slotting characteristic of elliptical wings. Sweepback and wide separation of the wing tips reduce “tip vortex,” a drag-creating turbulence that tends to develop at wing tips at faster speeds. This type of wing is aerodynamically efficient for high-speed flight but cannot easily keep a bird airborne at low speeds. The fastest birds, such as sandpipers, clocked at 175 km (109 miles) per hour, belong to this group.
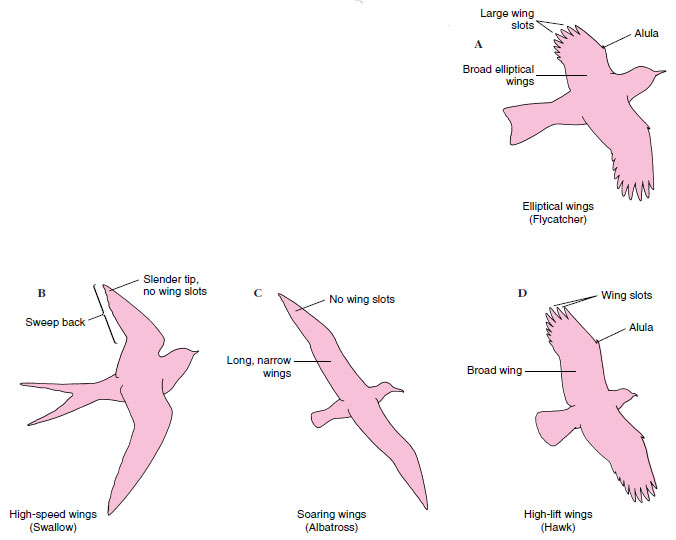 |
| Figure 29-19 Four basic forms of bird wings. |
Soaring Wings: The oceanic soaring birds have high-aspect ratio wings resembling those of sailplanes. This group includes albatrosses, frigate birds, and gannets (Figure 29-19C). Such long, narrow wings lack wing slots and are adapted for high speed, high lift, and dynamic soaring. They have the highest aerodynamic efficiency of all wings but are less maneuverable than the wide, slotted wings of land soarers. Dynamic soarers exploit the highly reliable sea winds, using adjacent air currents of different velocities.
High-Lift Wings: Vultures, hawks, eagles, owls, and ospreys (Figure 29-19D)—predators that carry heavy loads—have wings with slotting, alulas, and pronounced camber, all of which promote high lift at low speed. Many of these birds are land soarers, with broad, slotted wings that provide the sensitive response and maneuverability required for static soaring in capricious air currents over land.
Support our developers




