Origin and Relationships
Origin and
Relationships
Approximately 147 million years ago, a flying animal drowned and settled to the bottom of a shallow marine lagoon in what is now Bavaria, Germany. It was rapidly covered with a fine silt and eventually fossilized. There it remained until discovered in 1861 by a workman splitting slate in a limestone quarry. The fossil was approximately the size of a crow, with a skull not unlike that of modern birds except that the beaklike jaws bore small bony teeth set in sockets like those of reptiles (Figure 29-1). The skeleton was decidedly reptilian with a long bony tail, clawed fingers, and abdominal ribs. It might have been classified as a reptile except that it carried the unmistakable imprint of feathers, those marvels of biological engineering that only birds possess. Archaeopteryx lithographica (ar-kee-op´ter-ix lith-o-graf´e-ca, Gr., meaning “ancient wing inscribed in stone”), as the fossil was named, was an especially fortunate discovery because the fossil record of birds is disappointingly meager. The finding was also dramatic because it proved beyond reasonable doubt the phylogenetic relatedness of birds and reptiles.
Zoologists had long recognized the similarity of birds and reptiles. The skulls of birds and reptiles abut against the first neck vertebra by a single balland- socket joint, the occipital condyle (mammals have two condyles). Birds and reptiles have a single middle ear bone, the stapes (mammals have three middle ear bones). Birds and reptiles have a lower jaw composed of five or six bones, whereas the lower jaw of mammals has one mandibular bone, the dentary. Birds and reptiles excrete their nitrogenous wastes as uric acid whereas mammals excrete theirs as urea. Birds and reptiles lay similar yolked eggs with the early embryo developing on the surface by shallow cleavage divisions.
The distinguished English zoologist Thomas Henry Huxley was so impressed with these and many other anatomical and physiological affinites that he called birds “glorified reptiles” and classified them with a group of dinosaurs called theropods that displayed several birdlike characteristics (Figures 29-2 and 29-3). Theropod dinosaurs share many derived characters with birds, the most obvious of which is the elongate, mobile, S-shaped neck. As shown in the cladogram (Figure 29-3), theropods belong to a lineage of diapsid reptiles, the archosaurians, that includes crocodilians and pterosaurs, as well as the dinosaurs. There is now overwhelming evidence that Huxley was correct: birds’ closest phylogenetic affinity is to the theropod dinosaurs. The only anatomical feature required to link bird ancestry with the theropod dinosaurs was feathers, and this was provided by the discovery of Archaeopteryx. Recent discoveries of Cretaceous bird fossils in Spain, Madagascar, and China provide new data on bird ancestry. All these new discoveries, however, still link early birds with theropod reptiles.
Living birds (Neornithes) are
divided into two groups: (1) Paleognathae
(Gr. palaios, ancient, + gnathos, jaw), the large flightless
ostrichlike birds and the kiwis, often
called ratite birds, which have a flat
sternum with poorly developed pectoral
muscles, and (2) Neognathae (Gr. neos, new, + gnathos, jaw), flying birds
that have a keeled sternum on which
powerful flight muscles insert. This division
originated from the view that
flightless birds (ostrich, emu, kiwi, rhea)
represented a separate line of descent
that never attained flight. This idea is
now completely rejected. Ostrichlike paleognathids clearly have descended
from flying ancestors. Furthermore, not
all neognathous birds can fly and many
of them even lack keels (Figure 29-4).
Flightlessness has appeared independently
among many groups of birds; the
fossil record reveals flightless wrens,
pigeons, parrots, cranes, ducks, auks,
and even a flightless owl. Penguins are
flightless although they use their wings
to “fly” through water. Flightlessness
almost always has evolved on
islands where few terrestrial predators
are found. Flightless birds living on continents
today are the large paleognathids
(ostrich, rhea, cassowary,
emu), which can run fast enough to
escape predators. The ostrich can run
70 km (42 miles) per hour, and claims
of speeds of 96 km (60 miles) per hour
have been made.
Characteristics of Class Aves
Approximately 147 million years ago, a flying animal drowned and settled to the bottom of a shallow marine lagoon in what is now Bavaria, Germany. It was rapidly covered with a fine silt and eventually fossilized. There it remained until discovered in 1861 by a workman splitting slate in a limestone quarry. The fossil was approximately the size of a crow, with a skull not unlike that of modern birds except that the beaklike jaws bore small bony teeth set in sockets like those of reptiles (Figure 29-1). The skeleton was decidedly reptilian with a long bony tail, clawed fingers, and abdominal ribs. It might have been classified as a reptile except that it carried the unmistakable imprint of feathers, those marvels of biological engineering that only birds possess. Archaeopteryx lithographica (ar-kee-op´ter-ix lith-o-graf´e-ca, Gr., meaning “ancient wing inscribed in stone”), as the fossil was named, was an especially fortunate discovery because the fossil record of birds is disappointingly meager. The finding was also dramatic because it proved beyond reasonable doubt the phylogenetic relatedness of birds and reptiles.
 |
| Figure 29-1 Archaeopteryx, a 147-million-year-old ancestor of modern birds. A, Cast of the second and most nearly perfect fossil of Archaeopteryx, which was discovered in a Bavarian stone quarry. Seven specimens of Archaeopteryx have been discovered, the most recent one in 1992. B, Reconstruction of Archaeopteryx. |
Zoologists had long recognized the similarity of birds and reptiles. The skulls of birds and reptiles abut against the first neck vertebra by a single balland- socket joint, the occipital condyle (mammals have two condyles). Birds and reptiles have a single middle ear bone, the stapes (mammals have three middle ear bones). Birds and reptiles have a lower jaw composed of five or six bones, whereas the lower jaw of mammals has one mandibular bone, the dentary. Birds and reptiles excrete their nitrogenous wastes as uric acid whereas mammals excrete theirs as urea. Birds and reptiles lay similar yolked eggs with the early embryo developing on the surface by shallow cleavage divisions.
The distinguished English zoologist Thomas Henry Huxley was so impressed with these and many other anatomical and physiological affinites that he called birds “glorified reptiles” and classified them with a group of dinosaurs called theropods that displayed several birdlike characteristics (Figures 29-2 and 29-3). Theropod dinosaurs share many derived characters with birds, the most obvious of which is the elongate, mobile, S-shaped neck. As shown in the cladogram (Figure 29-3), theropods belong to a lineage of diapsid reptiles, the archosaurians, that includes crocodilians and pterosaurs, as well as the dinosaurs. There is now overwhelming evidence that Huxley was correct: birds’ closest phylogenetic affinity is to the theropod dinosaurs. The only anatomical feature required to link bird ancestry with the theropod dinosaurs was feathers, and this was provided by the discovery of Archaeopteryx. Recent discoveries of Cretaceous bird fossils in Spain, Madagascar, and China provide new data on bird ancestry. All these new discoveries, however, still link early birds with theropod reptiles.
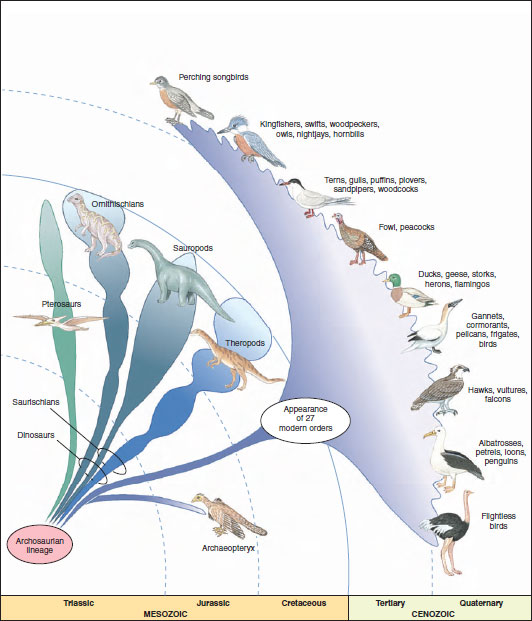 |
| Figure 29-2 Evolution of modern birds. Of 27 living bird orders, 9 of the largest are shown. The earliest known bird, Archaeopteryx, lived in the Upper Jurassic, about 147 million years ago. Archaeopteryx uniquely shares many specialized aspects of its skeleton with the smaller theropod dinosaurs and is considered to have evolved within the theropod lineage. Evolution of modern bird orders occurred rapidly during the Cretaceous and early Tertiary periods. |
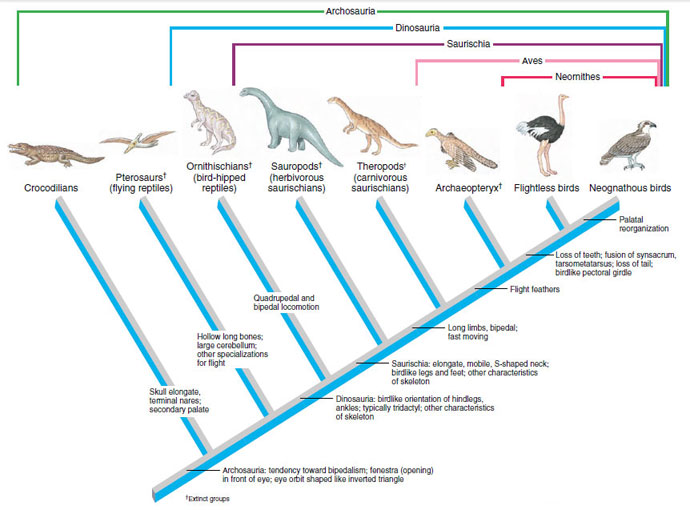 |
| Figure 29-3 Cladogram of the Archosauria, showing the relationships of several archosaurian groups to modern birds. Shown are a few of the shared derived characters, mostly those related to flight, that were used to construct the genealogy. The outgroup is Lepidosauria (see Figure 28-2) |
 |
| Figure 29-4 One of the strangest birds in a strange land, the flightless cormorant, Nannopterum harrisi, of the Galápagos Islands dries its wings after a fishing forage. It is a superb swimmer, propelling itself through the water with its feet to catch fish and octopuses. The flightless cormorant is an example of a carinate bird (having a keeled sternum) that has lost the keel and the ability to fly. |
Characteristics of Class Aves
- Body usually spindle shaped, with four divisions: head, neck, trunk, and tail; neck disproportionately long for balancing and food gathering
- Limbs paired with the forelimbs usually modified for flying; posterior pair variously adapted for perching, walking, or swimming; foot with four toes (2 or 3 toes in some)
- Epidermal covering of feathers and leg scales; thin integument of epidermis and dermis; no sweat glands; oil or preen gland at base of tail; pinna of ear rudimentary
- Fully ossified skeleton with air cavities; skull bones fused with one occipital condyle; skull diapsid with antorbital fenestra; each jaw covered with a horny sheath, forming a beak; no teeth; ribs with strengthening processes, the uncinate process attaching ribs with one another; tail not elongate; sternum well developed with keel or reduced with no keel; single bone in middle ear
- Nervous system well developed, with brain and 12 pairs of cranial nerves
- Circulatory system of fourchambered heart, with the right aortic arch persisting as the dorsal aorta; reduced renal portal system; nucleated red blood cells
- Endothermic
- Respiration by slightly expansible lungs, with thin air sacs among the visceral organs and skeleton; syrinx (voice box) near junction of trachea and bronchi
- Excretory system of metanephric kidney; ureters open into cloaca; no bladder; semisolid urine; uric acid main nitrogenous waste
- Sexes separate; testes paired, with the vas deferens opening into the cloaca; females with left ovary and oviduct only; copulatory organ in ducks, geese, paleognathids, and a few others
- Fertilization internal; amniotic eggs with much yolk and hard calcareous shells; embryonic membranes in egg during development; incubation external; young active at hatching (precocial) or helpless and naked (altricial); sex determination by females (females heterogametic)
Support our developers




