The gene transfer can be discussed using a hypothetical example of recombination between two auxotrophs. Let one of the auxotrophs be
methionineless (
met-)which will grow on a medium supplemented with methionine and let the other auxotroph be
threonineless (
thr-)which will grow on a medium supplemented with threonine. These two auxotrophs, can then be designated as
met-thr+and
met+thr-respectively. When these two auxotrophs are kept mixed for some time and the mixture spread on a minimal medium in an agar plate, a few colonies are formed. The colonies formed should be
met+thr+, because neither
met-thr+nor
met+thr-can grow on the minimal medium. Obviously due to recombination between
met-thr+ and
met+thr-, met+thr+ must have been produced from a zygote
met-thr+/
met+thr-.
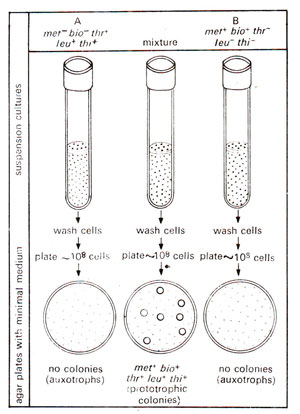
Fig. 12.3. Demonstration of genetic recombination between bacterial cells as demonstrated by Lederberg and Tatum (note the presence of colonies only when the mixture of A and B is used for plating).
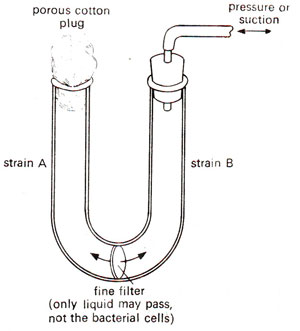
Fig. 12.4. Experiment demonstrating the need of physical contact between bacteria, for genetic recombination to occur (see text for details).
Lederberg and
Tatum, in their own experiment, utilized two strains of
Escherichia coli with different nutritional requirements : (i) strain A could grow on a minimal medium supplemented with methionine and biotin - designated as
met-bio-thr+leu+thi+ and (ii) strain B could grow on a minimal medium supplemented with threonine, leucine and thiamine - designated as
met+bio+thr-leu-thi-. Among petri-plates, all with unsupplemented minimal medium, some plates were plated with strain A, others with strain B and still others with a mixture of strain A and strain B, which had already been grown in a liquid medium containing all the supplements (Fig. 12.3). The plates were incubated overnight and the following observation were made : (i) No colonies were formed on plates incubated either with strain A or with strain B. (ii) Few colonies (1 for every 10,000,000 cells plated i.e. with a frequency of 1 x 10
-7) were observed on plates incubated with a mixture of both the strains. This suggested that the prototrophs
(met+bio+thr+leu+thi+) appeared due to recombination at a frequency of 1 x 10
-7.
Fig. 12.3. Demonstration of genetic recombination between bacterial cells as demonstrated by Lederberg and Tatum (note the presence of colonies only when the mixture of A and B is used for plating).
Two questions on the above experiment could be raised : (i) Could the wild type colonies be produced due to mutations ?, (ii) Could the growth of colonies be attributed to
'cross feeding' (leakage from one strain, of substances, which could be used by the other strain for growth) rather than genetic recombination. The first question could be answered, because if wild type colonies were due to mutation, these should have appeared even on plates incubated with individual strains. Further, the strain being deficient for two or three genes, the possibility of simultaneous mutation in two or three genes would be very remote and therefore, could be ruled out. For the answer to the second question, another experiment was conducted, where a U tube was used, with such a filter that the pores were too small to allow the bacteria to pass, but will be large enough to allow the passage of fluid with the dissolved substances.
Strain A was kept in one arm and strain B in the other arm (Fig. 12.4). After they had been incubated for a few hours, the contents of each arm were separately tested on minimal medium. None of them was found to grow on a minimal medium, suggesting that physical contact between strain A and strain B was necessary for the generation of wild type strains or prototrophs. In other words, gene transfer or genetic recombination involving physical contact was responsible for the appearance of prototrophs.
Fig. 12.4. Experiment demonstrating the need of physical contact between bacteria, for genetic recombination to occur (see text for details).




