Donor and recipient strains
Among the above strain A (met-bio-thr+leu+thi+) and strain B (met+bio+thr-leu-thi-), undergoing recombination, it was possible to designate them as donor and recipient strains on the basis of the following simple experiments involving treatment of either A or B with streptomycin (which does not kill immediately, but prevents cell division). When strain A was treated with streptomycin and mixed with strain B after washing out streptomycin, the frequency of colonies on minimal medium equaled those in control, where both strains were untreated. On the other hand,
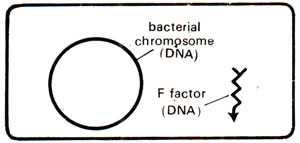
Fig. 12.5. A bacterial cell showing circular DNA and F factor.
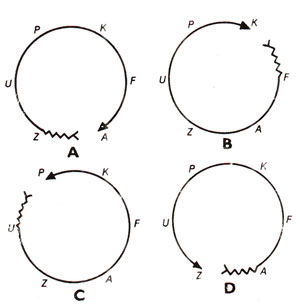
Fig. 12.6. (A, B, C, D). Four hypothetical Hfr strains having different genes at their proximal ends.
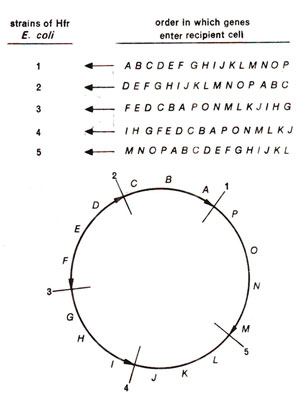
Fig. 12.7. The linear order of genes in five hypothetical Hfr strains, resulting due to breakage of bacterial chromosome at different sites and subsequent orientation acquired by the resulting linear chromosome
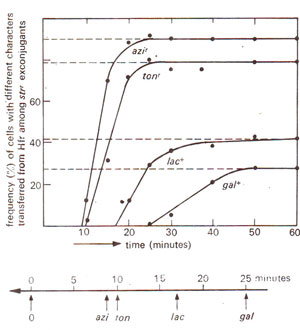
Fig. 12.8. (a) Results of a cross sirr, azis, tons, lac-, gal-(F ) x strs, azir, tonr, lac+, gal+ (Hfr), where samples from incubated mixture were drawn at different time intervals, disrupted in a blender, and spread on a medium containing streptomycin. The resistant colonies (sirr) were derived from F- parent (due to transfer of genes from Hfr) and were tested for four metabolic steps, (b) Chromosome map prepared from results shown in (a).

Fig. 12.9. Interconvefsion of F+ and Hfr strains, due to integration and detachment of F factor.
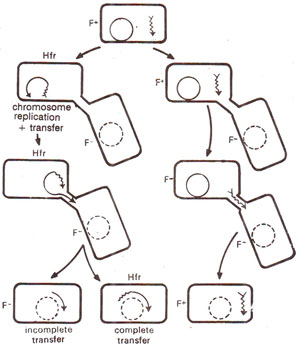
Fig. 12.10. Sexual conjugation, showing two alternatives. In one case (right) the F factor remains free, while in the other case (left) the F factor gets attached to form Hfr.
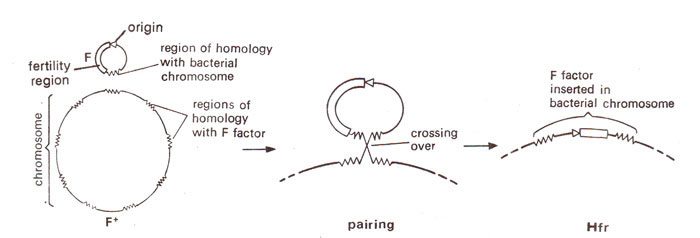
Fig. 12.11. A model for insertion of F factor into E. coli chromosome; an area of fertility factor has several regions of homology in E. coli chromosome, permitting crossing over (regions of homology are represented by jagged lines).
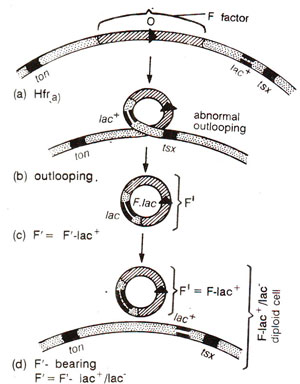
Fig. 12.12. Origin of F' factor (or F-lac) due to abnormal oullooping (a-c) and reintegration of F-lac+ into F-lac-strain.
Fig. 12.9. Interconvefsion of F+ and Hfr strains, due to integration and detachment of F factor.
When conjugation occurs between F+ donor and F- recipient, two possibilities exist (Fig. 12.10) (i) F factor may remain free in cytoplasm, so that only F factor will enter the recipient cell converting the recipient F- into a donor F+. (ii) F factor may get integrated and would thus make the distal part of bacterial chromosome. In this case the chromosome will become linear and would have a directional orientation, so that it would enter the recipient cell from the end away from attached F factor.
Fig. 12.10. Sexual conjugation, showing two alternatives. In one case (right) the F factor remains free, while in the other case (left) the F factor gets attached to form Hfr.
From the above discussion, it also emerges that the low recombination attribute of F+ strain is due to presence of Hfr among F+ cells in low frequency. When same Hfr is found in a relatively pure state, recombination frequency increases. In other words, for recombination to take place, F factor must be attached to bacterial chromosome and the circular chromosome should take up a linear orientation.
Conversion of F+ into Hfr strain
It has been shown that like the bacterial chromosome, fertility factor F is also circular, so that a crossover between the two rings would produce a larger ring with F inserted. The F is believed to consist of three regions : (i) origin, (ii) fertility genes and (iii) pairing region. The pairing region of F has homology with several regions of bacterial chromosome, so thatcrossing over can take place at any one of these regions, producing different Hfr strains (Fig. 12.11).
Fig. 12.11. A model for insertion of F factor into E. coli chromosome; an area of fertility factor has several regions of homology in E. coli chromosome, permitting crossing over (regions of homology are represented by jagged lines).
Fertility factor F' and sexduction
During experiments with Hfr strains, initially (in 1959) an Hfr strain was found, which kept on producing F+ cells at high frequencies. The fertility factor involved in this process was described as F' to distinguish it from normal F,
Fig. 12.12. Origin of F' factor (or F-lac) due to abnormal oullooping (a-c) and reintegration of F-lac+ into F-lac-strain.
Support our developers




