Application of Potassium Fertilizers
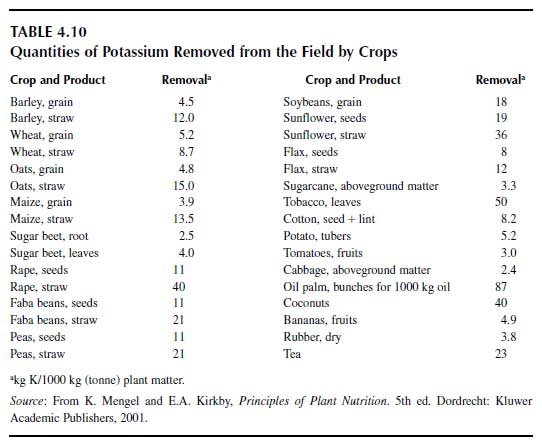
Chlorophobic crop species should not be fertilized with potassium chloride. Such species are tobacco (Nicotiana tabacum L.), grape (Vitis vinifera L.), fruit trees, cotton, sugarcane (Saccharum officinarum L.), potato, tomato, strawberry (Fragaria x ananassa Duchesne), cucumber (Cucumis sativus L.), and onion (Allium cepa L.). These crops should be fertilized with potassium sulfate. If potassium chloride is applied, it should be applied in autumn on soils that contain sufficiently high concentrations of K+-selective binding sites in the rooting zone. In such a case, the chloride may be leached by winter rainfall, whereas the K+ is adsorbed to 2:1 minerals and hence is available to the crop in the following season. On soils with a medium to high cation exchange capacity (CEC>120 mgmol/kg) and with 2:1 selective K+-binding minerals, potassium fertilizers can be applied in all seasons around the year since there is no danger of K+ leaching out of the rooting profile (Alfisols, Inceptisols, Vertisols, and Mollisols, in contrast to Ultisols, Oxisols, Spodosols, and Histosols). In the latter soils, high K+ leaching occurs during winter or monsoon rainfall. Histosols may have a high CEC on a weight basis but not on a volume basis because of their high organic matter content. In addition, Histosols contain few K+-selective binding sites. Under tropical conditions on highly weathered soils (Oxisols, Ultisols), potassium fertilizer may be applied in several small doses during vegetative growth in order to avoid major K+ leaching.The quantities of fertilizer potassium required depend on the status of available K+ in the soil and on the crop species, including its yield level. Provided that the status of available K+ in the soil is sufficient, the potassium fertilizer rate should be at least as high as the quantity of potassium present in the crop parts removed from the field, which in many case are grains, seeds, tubers, roots or fruits. In Table 4.10 (15), the approximate concentrations of potassium in plant parts are shown. It is evident that the potassium concentrations in cereal grains are low compared with leguminous seeds, sunflower (Helianthus annuus L.) and rape seed. Potassium removal by fruit trees is shown in Table 4.11. The concept of assessing fertilizer rates derived from potassium removal is correct provided that no major leaching losses occur during rainy seasons. In such cases, the K+ originating from leaves and straw remaining on the field may be leached into the subsoil at high rates. Such losses by leaching are the case for Spodosols, Oxisols, and Ultisols. Here, besides the K+ removed from the soil by crop plants, the leached K+ must also be taken into consideration.
On the other hand, if a soil has a high status of available K+, one or even several potassium fertilizer applications per crop species in the rotation may be omitted. As a first approach for calculating the amount of available K+ in the soil, 1 mg/kg soil of exchangeable K+ equals approximately 5 kg K/ha. In this calculation, interlayer K+ is not taken into consideration. If the soil is low in available K+, for most soils higher fertilization rates are required than 5 kg K/ha per mg exchangeable K+, since with the exception of Histosols and Spodosols, sites of interlayer positions must be filled up by K+ before the exchangeable K+ will be raised. This problem is particularly acute on K+-fixing soils. Here, high K fertilizer rates are required, as shown in Table 4.12 (112). From the discussion, it is clear that with normal potassium fertilizer rates, the yield and the potassium concentration in leaves were hardly raised and optimum yield and leaf potassium concentrations were attained with application of 1580 kg K/ha. As soon as the K+-fixing binding sites are saturated by K+, fertilizer should be applied at a rate in the range of the K+ accumulation by the crop.
 |
 |
Plant species differ in their capability for exploiting soil K+. There is a major difference between monocotyledonous and dicotyledonous species, the latter being less capable of exploiting soil K+, mainly interlayer K+, than the former. In a 20-year field trial on an arable soil derived from loess (Alfisol), the treatment without potassium fertilizer produced cereal yields that were not much lower than those in the fertilized treatment, in contrast to the yields of potatoes, faba beans (Vicia faba L.), and a clover-grass mixture. With these crops, the relative yields were 73, 52, and 84, respectively, with a yield of 100 in the potassium-fertilized treatment (113). This different behavior is particularly true for grasses and leguminous species. Root investigations under field conditions with perennial ryegrass and red clover (Trifolium pratense L.) cultivated on an Alfisol showed considerable differences in root morphology, including root hairs and root length, which were much longer for the grass (114). Hence the root-soil contact is much greater for the grass than for the clover. The grass will therefore still feed sufficiently from the low soil solution K+ concentration originating from interlayer K+, a concentration that is insufficient for the clover. From this result, it follows that leguminous species in a mixed crop stand, including swards of meadow and pasture, will withstand the competition with grasses only if the soil is well supplied with available K+.
 |
This difference between monocots and dicots in exploiting soil K+ implies that grasses can be grown satisfactorily on a lower level of exchangeable soil K+ than dicots. It should be taken into consideration, however, that a major depletion of interlayer K+ leads to a loss of selective K+-binding sites because of the conversion or destruction of soil minerals (92), giving an irreversible loss of an essential soil fertility component.
Table 4.12 shows that the optimum K+ supply considerably decreases the percentage of crop lodging. This action is an additional positive effect of K+, which is also true with other cereal crops. As already considered above, K+ favors the energy status of plants and thus the synthesis of various biochemical compounds such as cellulose, lignin, vitamins, and lipids. In this respect, the synthesis of leaf cuticles is of particular interest (15). Poorly developed cuticles and also thin cell walls favor penetration and infection by fungi and lower the resistance to diseases (115).
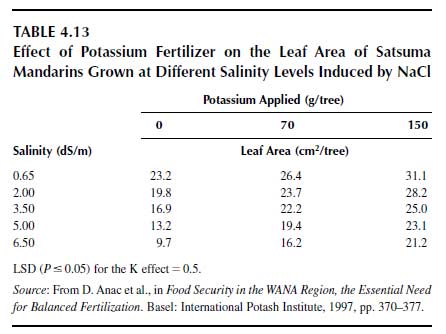
Heavy potassium fertilizer rates also may depress the negative effect of salinity since the excessive uptake of Na+ into the plant cell is depressed by K+. Table 4.13 presents such an example for mandarin oranges (Citrus reticulata Blanco) (116), showing that the depressive effect of salinity on leaf area was counterbalanced by higher potassium fertilizer rates. The higher the relative K+ effect, the higher is the salinity level.
 |
Support our developers




