Interactions between Sulfur and Other Minerals
Interactions between sulfur and other minerals may significantly influence crop quality parameters (5,113,114). Sulfur and nitrogen show strong interactions in their nutritional effects on crop growth and quality due to their mutual occurrence in amino acids and proteins (see Section).Nitrogen-Sulfur Interactions
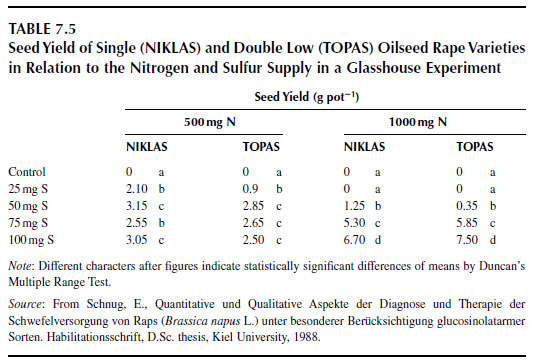
Under conditions of sulfur starvation, sulfur deficiency symptoms are expressed moderately at low nitrogen levels but extremely with a high nitrogen supply. This effect explains the enhancement of sulfur deficiency symptoms in the field after nitrogen dressings (114). The question of why sulfur deficiency symptoms are more pronounced at high nitrogen levels is, however, still unanswered. For experimentation, these results are relevant as the adjustment of the nitrogen and sulfur nutritional status of plants is essential before any hypothesis on the effect of a nitrogen or sulfur treatment on plant parameters can be stated or proved.The use of the nitrogen/sulfur ratio as a diagnostic criterion is problematic because the same ratio can be obtained at totally different concentration levels in the tissue. Surplus of one element may therefore be interpreted falsely as a deficiency of the other (see Section). Clear relationships between nitrogen/sulfur ratios and yield occur only in ranges of extreme ratios. Such ratios may be produced in pot trials but do not occur under field conditions. The effect of increasing nitrogen and sulfur supply on crop seed yield with increasing nitrogen supply is more pronounced with protein than with carbohydrate crops (Table 7.5).
 |
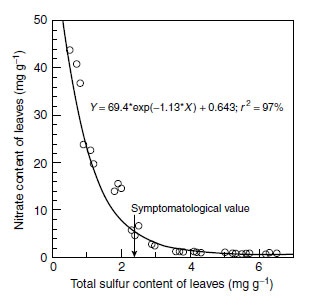 |
| FIGURE 7.6 Nitrate concentrations in the dry matter of lettuce in relation to the sulfur nutritional status of the plants. (From Schnug, E., in Sulphur Metabolism in Higher Plants: Molecular, Ecophysiological and Nutritional Aspects, Backhuys Publishers, Leiden, 1997, pp. 109-130.) |
Changes in the nitrogen supply affect the sulfur demand of plants and vice versa. Under conditions of sulfur deficiency, the utilization of nitrogen will be reduced and consequently nonprotein nitrogen compounds, including nitrate, accumulate in the plant tissue (Figure 7.6) (5,112).
The antagonistic relationship between sulfur supply and nitrate content exists in the range of severe sulfur deficiency, when macroscopic symptoms are visible. The higher the nitrogen level in the plants, the stronger the effect on the nitrate content will be. Thus, an adequate sulfur supply is vital for minimizing undesired enrichment with nitrate.
Photosynthesis and growth of pecan (Carya illinoinensis Koch) increased with N supply in relation to the nitrogen/sulfur ratio in pecan leaves (115). Both parameters were, however, reduced when combined leaf nitrogen and sulfur concentrations of <35 mg g-1 nitrogen and 3.7 mg g-1 sulfur were noted (115).
The initial supply of a crop with nitrogen and sulfur is decisive for its influence on the glucosinolate content, probably due to physiological or root-morphological reasons (103). Nitrogen fertilization to oilseed rape insufficiently supplied with nitrogen and sulfur will lead to decreasing glucosinolate concentrations because the demand of an increasing sink due to increasing numbers of seeds will not be met by the limited sulfur source. Only if the rooting depth or density is enhanced by the nitrogen supply, which increases the plant-available sulfur pool in the soil, does the glucosinolate content increase too. Higher glucosinolate concentrations in seeds can also be expected after nitrogen applications to crops with a demand for nitrogen but adequate sulfur supply due to the increased biosynthesis of sulfur-containing amino acids, which are precursors of glucosinolates. In the case of a crop already sufficiently supplied with nitrogen, there is no evidence for any specific nitrogen-sulfur interactions on the glucosinolate content (5,116).
In general, no significant influence of nitrogen fertilization on the alliin content has been found for onions (Allium cepa L.) and garlic (Allium sativum L.), but there is a tendency that a higher nitrogen supply results in a decreased alliin content (101). In comparison, an increasing sulfur supply has been related to an increasing alliin content in leaves and bulbs of both crops. There were also interactions between nitrogen and sulfur in such a way that the total sulfur content of onion leaves was correlated highly with nitrogen fertilization: the sulfur concentration of leaves decreased with increasing N fertilization, and the total nitrogen concentration of onion bulbs decreased with increasing sulfur fertilization. The same observations were made by Freeman and Mossadeghi (117) for garlic plants, where the nitrogen concentration decreased from 4.05 to 2.93% with sulfur fertilization, and by Randle et al. (118), who reported decreasing total bulb sulfur concentrations in response to increasing nitrogen fertilization.
Interactions between Sulfur and Micronutrients
Owing to antagonistic effects, sulfur fertilization reduces the uptake of boron and molybdenum. In soils with a marginal plant-available concentration of these two plant nutrients, sulfur fertilization may induce boron or molybdenum deficiency, particularly on coarse-textured sites where brassica crops are grown intensely in the crop rotation (119). In comparison, sulfur fertilization is an efficient tool to reduce the selenium, molybdenum, arsenic, bromine, and antimony uptake on contaminated sites. The influence of elemental sulfur applications on the concentration of trace elements of fully developed leaves of nasturtium (Tropaeolum majus L.) was tested on two sites in northern Germany (120). The results of this study reveal a significantly increased uptake of copper, manganese, cobalt, nickel, and cadmium, with increasing levels of sulfur. This increased uptake was caused by a higher availability of these elements due to the acidifying effect of elemental sulfur. At the same time, antagonistic effects were noted for arsenic, boron, selenium, and molybdenum in relation to the soil type.The enzyme sulfite oxidase is a molybdo-enzyme, which converts sulfite into sulfate (121) and is thus important for sulfate reduction and assimilation in plants (see Figure 7.1). Stout and Meagher (122) have shown that the sulfate supply influences molybdenum uptake. Sulfate-molybdate antagonism can be observed at the soil-root interface and within the plant, as an increasing sulfur supply results in lower molybdenum concentrations in the tissues (123). The significance of sulfate-molybdate antagonism in agriculture is described comprehensively by Macleod et al. (124).
Selenium, like molybdenum, is chemically similar to sulfur. Comprehensive reviews about interactions between sulfate transporters and sulfur assimilation enzymes, and selenium-molybdenum uptake and metabolism, are given by Terry et al. (125) and Kaiser et al. (126). Accumulation of glutathione due to elevated levels of sulfate in the soil and SO2/H2S in the air was reduced drastically in spinach (Spinacia oleracea L.) leaf discs by selenate amendments (127). In those studies the uptake of sulfur was not influenced by the selenate treatment. Bosma et al. (128) suggested that selenate decreases sulfate reduction due to antagonistic effects during plant uptake, in combination with a rapid turnover of glutathione. An increasing sulfate supply gives higher sulfate concentrations in the plant tissue, so that the competition between sulfur and selenium for the enzymes of the sulfur assimilation pathway will finally result in less synthesis of selenoamino acids (129).
This antagonistic effect is of no practical significance on seleniferous soils, but it could be relevant on deficient and marginal sites (130). Field experiments with combined sulfur and selenium applications to grass-clover pastures, on selenium-deficient and high-selenium sites revealed that selenium concentrations in the different botanical species showed distinct differences in relation to the site (130).
On the high-selenium site, sulfur fertilization significantly decreased the selenium concentration in pasture. Spencer (130) attributed this action to a dilution effect, as the total selenium content remained constant. Studies on the pungency of onion bulbs in relation to the sulfur supply revealed that although sulfur content was increased at elevated selenium levels, the pungency was reduced (131). Kopsell and Randell (131) proposed that selenium had an impact on the biosynthetic pathway of flavor precursors.
A synergistic effect of sulfur and selenium on the shoot sulfur concentration was noted for hydroponically grown barley (Hordeum vulgare L.) and rice (Oryza sativa L.). With increasing selenium concentrations in the solution, a steep increase in the sulfur concentration of the shoots occurred even with a low sulfur supply (132).
Sulfur and phosphorus interactions in plants are closely related to plant species, because of the different root morphologies and nutrient demands of different species (133). A synergistic effect of sulfur and phosphorus on crop yield occurred for sorghum (Sorghum vulgare Pers.), maize (Zea mays L.), wheat (Triticum aestivum L.), and mustard (Brassica spp. L.) (134-137). A synergistic relationship between sulfur and potassium, which enhances crop productivity and quality, was determined in several studies (138-140).
Support our developers




