Identification of Limiting Steps in the PCR Cycle
The primary reactions of photosynthesis can be roughly divided into four parts: formation of NADPH and ATP, incorporation of CO2 into ribulose 1,5-bisphosphate (RuBP) by RuBisCO to produce 3-phosphoglycerate (PGA), regeneration of RuBP in the PCR cycle (Fig. 4.1), and sucrose synthesis using triose phosphate exported into the cytosol and counter changed with phosphate released by this synthesis. The accepted photosynthesis model (Farquhar et al., 1981) is based on the prediction that the rate of synthesis of NADPH and ATP is calculated from the flux of electrons in the photosynthetic electron transport chain, with three protons transported for every ATP formed. In situ RuBisCO activity is calculated using the concentration of the activated catalytic site and kinetic parameters of RuBisCO (Farquhar, 1979). The steady-state concentration of RuBP is balanced both by the rate of regeneration and the utilization by RuBisCO for CO2 fixation. Important information has been provided by simultaneous measurements of rates of gas exchange and steady-state concentrations of metabolites in the PCR cycle using part of a single attached leaf under a range of conditions. The photosynthetic rate of an attached leaf has been found to match the rate calculated with RuBisCO kinetics at CO2 concentrations in the intercellular space below 40 Pa and at saturating light intensities, while the photosynthetic rate calculated by taking electron flux into consideration significantly exceeds the photosynthetic rate (Badger et al., 1984). The intraplastidic concentration of RuBP reaches levels that are several fold higher than the concentration of the RuBisCO active site under these conditions (Badger et al., 1984; Geiger and Servaites, 1994). This indicates that photosynthesis is limited by either RuBisCO or the CO2-fixation pathway. As the intercellular CO2 concentration increases, photosynthesis enters an RuBP-limited phase and transport of inorganic phosphate back into chloroplasts becomes rate limiting (Sage, 1990; Sage et al., 1989). In contrast, the capacity for NADPH and ATP formation limits photosynthesis at nonsaturating light intensities (Farquhar et al., 1981). Moreover, photosynthesis in source organs may occasionally become limited by the capacities of sink organs to accumulate photosynthates (Paul and Foyer, 2001).
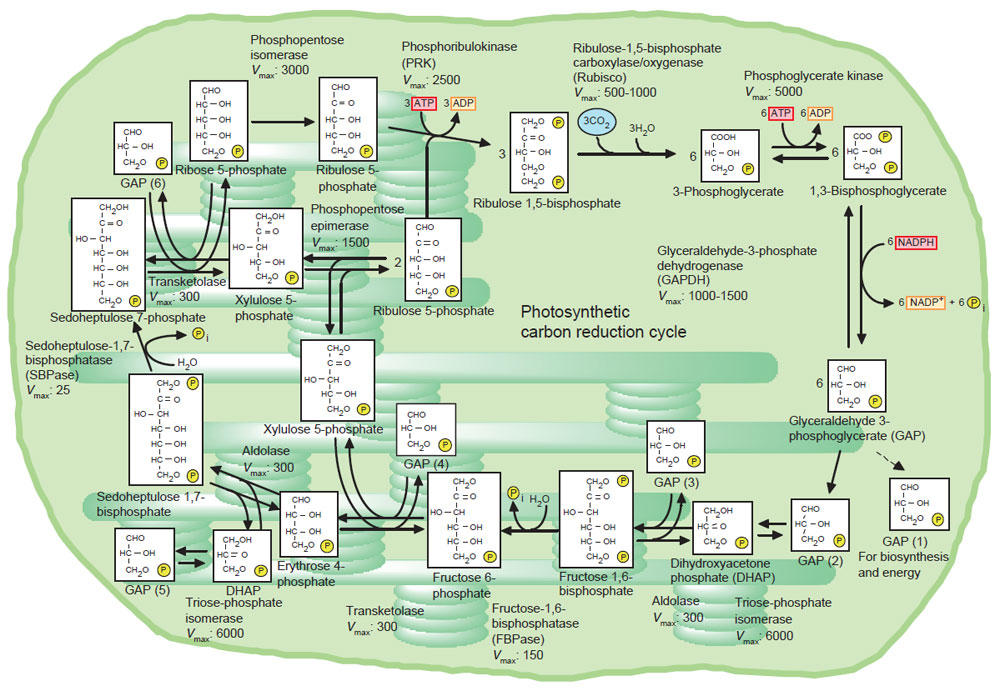 |
| FIGURE 4.1 Photosynthetic carbon reduction cycle. Vmax of each enzyme is given in micromoles per milligram chlorophyll per hour (Robinson and Walker, 1981). |
Support our developers




