Endothelial Cell Invasion Assay
I. INTRODUCTIONAngiogenesis is defined as the sprouting of new capillaries from preexisting blood vessels (Folkman and Shing, 1992). The formation of new blood vessels occurs in a variety of normal and pathologic conditions. During embryogenesis or wound healing, neovascularization is the result of a balance of stimulatory and inhibitory angiogenic factors. Normally this balance is strictly controlled. However, during the development of many diseases, including inflammation, retinopathies, and cancer metastasis, the angiogenesis controlling mechanisms may fail and result in formation of a pathologic capillary network (Folkman, 1992).
During vascular assembly, endothelial cells respond to a variety of extracellular growth factors and different molecules involved in cell-cell and cell-matrix interactions (Gale and Yancopoulos, 1999). Signaling molecules, more commonly associated with neuronal development, also play an important role in capillary formation during angiogenesis (Wang et al., 1998; Soker et al., 1998).
The angiogenic response is considered to be composed of a series of sequential steps, including degradation of the basement membrane surrounding the endothelial cells, migration of the endothelial cells into the surrounding tissues, proliferation and differentiation of the endothelial cells, and finally formation of new capillary vessels (Folkman, 1986). Most of these distinctive steps of the angiogenic process can be mimicked in vitro. in vitro assays classically rely on the use of two sources of endothelial cells: large vessel endothelial cells, such as bovine aortic endothelial cells (BAEC), or human umbilical vein endothelial cells (HUVEC) (Zetter, 1988; Springhorn et al., 1995), and endothelial cells of microvascular origin (Kråling et al., 1994; Lamszus et al., 1999). in vitro assays are quantifiable and may specifically address each step of the angiogenic cascade such as basement membrane disruption (metalloproteinase assays), cell migration (in vitro wounding; phagokinetic track assay; modified Boyden chamber chemotaxis assay), proliferation (thymidine incorporation assay), and tube formation by endothelial cells in three-dimensional gels, such as collagen or Matrigel. Matrigel is a tumor basement membrane matrix extract enriched with laminin. During angiogenesis, the proliferating and migrating endothelial cells are eventually organized into newly formed capillary structures. Endothelial cells of all origins are able to form tube-like structures in vitro when growing in appropriate extracellular matrix components. Matrigel considerably enhances tube formation (Grant et al., 1985; Madri et al., 1988).
There are several aspects of cell culture assays involving endothelial cells that need to be taken into account for the accurate interpretation of the obtained results. Prolonged cultivation of endothelial cells may change their properties dramatically, including alterations in activation state, karyotype, expression of cell surface antigens, and growth properties. Another problem is that all endothelial cells are not alike. Microvascular endothelial cells differ in different organs or even are different within blood vessels of the same organ (Gumkowski et al., 1991). Different endothelial cells produce different factors (cytokines, growth factors, and their inhibitors) that make the assay system even more complex for evaluation. From that point of view, organ culture assay systems including aortic ring assay (Nicosia and Ottinett, 1990) and chick aortic arch assay (Muthukkaruppan et al., 2000) are even more complex. In these assays, entire aortic segments, including nonendothelial cells, are placed in Matrigel. In 1-2 weeks the segments are monitored for the outgrowth of endothelial cells and for the formation of three-dimensional tube-like structures. The main drawback of this system is using adult aorta endothelial cells in the case of the aortic ring assay or microvascular endothelial cells obtained from embryonic arch, which are composed of cells dividing before exposure to angiogenic factors. Nevertheless the ability of endothelial cells to form tube-like structures in Matrigel makes this in vitro cell culture model the most faithful assay system.
This article describes the modification of a tube forming assay in Matrigel in which a dense clump of cultured endothelial cells is formed by cultivation of the cells in a hanging drop of growth media. During incubation, cells form dense "clump" at the bottom of the drop. The sediment of the cells is transferred onto a layer of Matrigel and is covered with a thin layer of Matrigel in which an interconnecting network of endothelial capillary tubes is formed rapidly. Tube formation occurs through an ordered sequence of events. Endothelial cells localized on the surface of the "cellular island" first develop large, dynamic cellular protrusions and then form small aggregates and cord-like structures. These early cord structures are dense and do not have lumens. Cells start to migrate to form a complex network of tube-like structures. The advantages of the assay are the ability to directly visualize the changes in cell morphology and the ability of endothelial cells to generate tube-like structures. Endothelial cells in the "clump" can survive for more than a few days, leaving adequate time for assessing angiogenic reactions. Introduction of angiogenesisinducing factors or cells into Matrigel at some distance from the "island" allows one to test chemotactic activity. The assay is readily quantifiable but utilizes already mentioned problems characteristic of the in vitro angiogenesis assay.
Another advantage is that in this assay, cultivated endothelial cell lines can be used, which can give more standard results than the use of primary cultures, or organ cultures.
II. MATERIALS AND INSTRUMENTATION
Growth medium: Dulbecco's modified Eagle's medium (DMEM) with a high concentration of glucose (Cat. No. 31966-021,GIBCO) supplemented with fetal bovine serum (FBS, Cat. No. F 7524, Sigma) at 10%, 100× penicillin/streptomycinx (Cat. No. 15140-148, GIBCO).
PBS(-) Ca2+ and Mg2+-free Dulbecco's phosphatebuffered saline (PBS, 2.7mM KH2PO4, 8.1mM Na2HPO4, 137 mM NaCl), sterile.
Trypsin-EDTA solution (Cat. No. 25200-056, GIBCO)
HEPES buffer solution 1M (Cat. No. 15630-049, GIBCO)
Stock of Matrigel - basement membrane matrix (BD Biosciences, Cat. No. 354234). Matrigel is supplied frozen and is stored at -70°C.
25-cm2 tissue culture flasks, 48-well tissue culture plates, Pasteur pipettes, 10-ml pipettes (sterile), and syringe needles (23 gauge).
III. PROCEDURES
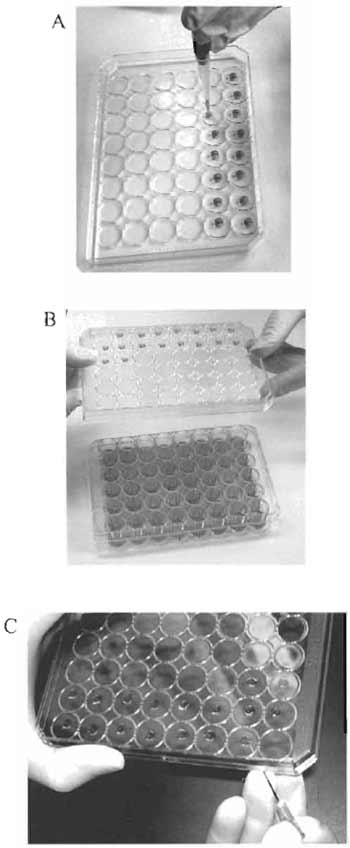 |
| FIGURE 1 Culturing cells in the hanging drops. (A) Plating of the cell suspension in the middle of the inner side of the lid. (B) Turning over the lid with the hanging drops. (C) Transferring the cell "clump" with a syringe needle. |
- For culture in 75-cm2 flasks, suspend murine microvascular endothelial SVEC4-10 cells in culture medium at a concentration of 2 × 105 cells/ml and plate out 2-4ml of cell suspension/flask. Place the flasks in a 37°C tissue culture incubator and incubate in an atmosphere of 5% CO2 overnight.
- The next day, examine cells under an inverted microscope fitted with phase-contrast objectives. Choose the flasks with the cell density >80% of confluence.
- Remove media and wash the cells with 10ml PBS(-).
- Replace the PBS with 1 ml of trypsin-EDTA and incubate at 37°C for approximately 1 min. Cell rounding should be observed in the inverted microscope. When the cells are rounded, detach them by strong agitation. Add 10 ml of culture medium supplemented with 10% FBS to the flask, pipette the cells up and down five times, and transfer contents to a 15-ml centrifuge tube.
- Centrifuge the cells at 800g for 5min at room temperature. Resuspend the cells in 10ml DMEM/ FBS. Count an aliquot of the cell suspension with the Coulter counter or in a hemocytometer.
- Centrifuge the cells at 800g for 5min at room temperature.
- Resuspend the cells in DMEM supplemented with 20% FBS, 1M HEPES, pH 7.5, at a concentration of 3 × 106 cells/ml.
- Dispense 1.0ml of DMEM/FBS into each well of a 48-well tissue culture plate.
- Turn the lid of the plate upside down. Plate out 0.02 ml of the cell suspension in the middle of the inner side of the lid. The cell suspension should form a drop (Fig. 1A).
- Carefully turn over the lid and place it on the plate. The drops of the cell suspension will hang over the media in the wells (Fig. 1B).
- Incubate plates at 37°C in a tissue culture incubator. During overnight incubation, cells form a dense "clump" at the bottom of the drop.
- Defreeze the necessary amount of Matrigel overnight at 4°C with rotation.
- The next day, examine all the wells under a stereomicroscope and select those that have the most compact, well-formed "clump" of cells.
- Place Matrigel on ice. Add 5 × DMEM and FBS to obtain a final concentration of 1× DMEM and 10% FBS. Keep the mixture on ice.
- Place a fresh 48-well plate on ice. Dispense 0.15ml/well of Matrigel/mix into the plate. Incubate the plate at 37°C in an incubator for 3-4 hours. The Matrigel should solidify.
- Carefully lift the lid of the plate with hanging drops, stick the cell "clump" to the tip of a syringe needle, and transfer the "clump" on the surface of the solidified Matrigel in the 48-well tissue culture plate (Fig. 1C).
- Cover the cell "clump" with 0.01 ml of Matrigel/ mix and incubate the plate for 15min at 37°C in an incubator.
- Add 0.3ml of DMEM/FBS in each well and return the plate to the incubator. The cells will remain viable for several days.
IV. COMMENTS
Using the protocol described in this assay, it is possible to demonstrate the effects of compounds on the ability of endothelial cell to form capillary-like tubes in vitro. We have also used this protocol to cocultivate endothelial cells with other cell types and to study the influence of the molecules produced by these cells on the ability of endothelial cells to form capillaries (Fig. 2D).
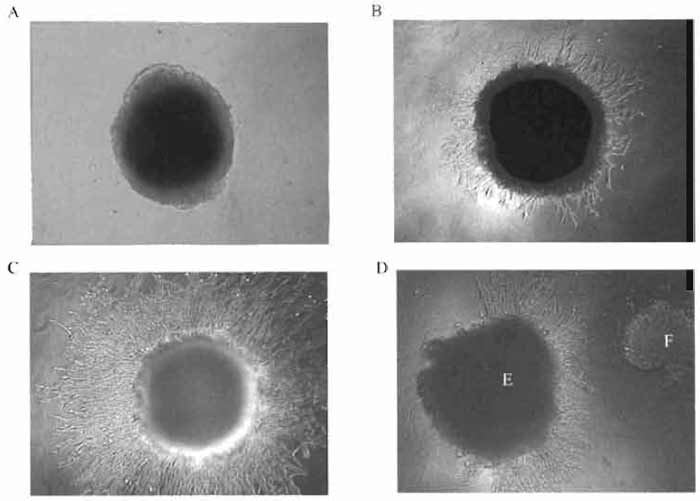 |
| FIGURE 2 Mouse endothelial cells SVEC4-10 migrating from a dense "clump" of the cells into Matrigel containing an angiogenic compound: (A) 0 h of cultivation, (B) 24h, (C) 48 h, and (D) chemotactic activity of an angiogenic compound. E, endothelial cells; F, tumor cells. |
V. PITFALLS
- To obtain maximum viability of cells growing in the hanging drops, avoid long exposure of the cells to trypsin during harvesting cells from the 75-cm2 flasks.
- Choose only well-formed, round-shaped, dense cellular "clumps."
- When lifting the lid with the hanging drops, caution must be exercised to avoid disrupting the cell "clumps."
References
Folkman, J. (1986). How is blood vessel growth regulated in normal and neoplastic tissue? G.H.A. Clowes memorial award lecture. Cancer Res. 46, 467-473.
Folkman, J. (1992). The role of angiogenesis in tumor growth. Semin. Cancer Biol. 3, 65-71.
Folkman, J., and Shing (1992). Angiogenesis. J.Biol.Chem. 267, 10931-1093.
Gale, N., and Yancopoulos, G. D. (1999). Growth factors acting via endothelial cell-specific receptor tyrosine kinases: VEGFs, angiopoietins and ephrins in vascular development. Genes Dev. 13, 1055-1066.
Grant, D. S., Kleinman, H. K., Leblong, C. P., Inoue, S., Chung, A. E., and Martin, G. R. (1985). The basement-membrane-like matrix of the mouse EHS tumor. II. Immunochemical quantitation of six of its components. Am.J.Anat. 174, 387-398.
Gumkowski, E, Kaminska, G., Kaminski, M., Morrissey, L. M., and Auerbach R. (1991). Heterogeneity of mouse vascular endothelium: in vitro studies of lymphatic, large blood vessels and microvascular endothelial cells. Blood Vessels 24, 11-23.
Kråling, B. M., Jimenez, S. A., Sorger, T., and Maul, G. G. (1994). Isolation and characterization of microvascular endothelial cells from the adult human dermis and from skin biopsies of patients with systemic sclerosis. Lab. Invest. 71, 745-754.
Lamszus, K., Schmidt, N. O., Ergun, S., and Westphal, M. (1999). Isolation and culture of human neuromicrovascular endothelial cells for the study of angiogenesis in vitro. J. Nerosci. Res. 55, 370-381.
Madri, J. A., Pratt, B. M., and Tucker, A. M. (1988). Phenotipic modulation of endothelial cells by transforming growth factorq3 depends upon the composition and organization of the extracellular matrix. J. Cell Biol. 106, 1375-1384.
Muthukkaruppan, V. R., Shinners, B. L., Lewis, R., Park, S-J., Baechler, B. J., and Auerbach, R. (2000). The chick embryo aortic arch assay: A new, rapid, quantifiable in vitro method for testing the efficacy of angiogenic and anti-angiogenic factors in a threedimensional, serum-free organ culture system. Proc. Am. Assoc. Cancer Res. 41, 65.
Nicosia, R. E, and Ottinett, A. (1990). Growth of microvessels in serum-free matrix culture of rat aorta. A quantitative assay of angiogenesis in vitro. Lab Invest. 63, 115-122.
Soker, S., Takashima, S., Miao, H. Q., Neufeld, G., and Klagsbrun, M. (1998). Neuropilinl is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 92, 735-745.
Springhorn, J. P., Madri, J. A., and Squinto, S. P. (1995). Human capillary endothelial cells from abdominal wall adipose tissue: Isolation using an anti-PECAM antibody. in vitro Cell Dev. Biol 31, 473-481.
Wang, H. U., Chen, Z. E, and Anderson, D. J. (1998). Molecular distinction and angiogenic interaction between embryonic arteries and viens revealed by ephrin B2 and its receptor EphB4. Cell 93, 741-753.
Zetter, B. R. (1988). Endothelial heterogeneity: Influence of vessel size, organ localization, and species specificity on the properties of cultured endothelial cells. In "Endothelial Cells" (U.S. Ryan, ed.), vol. 2, pp. 63-79. V. 2. CRC, Boca Raton, FL.
Support our developers




