Vitamins and diet enrichment compounds
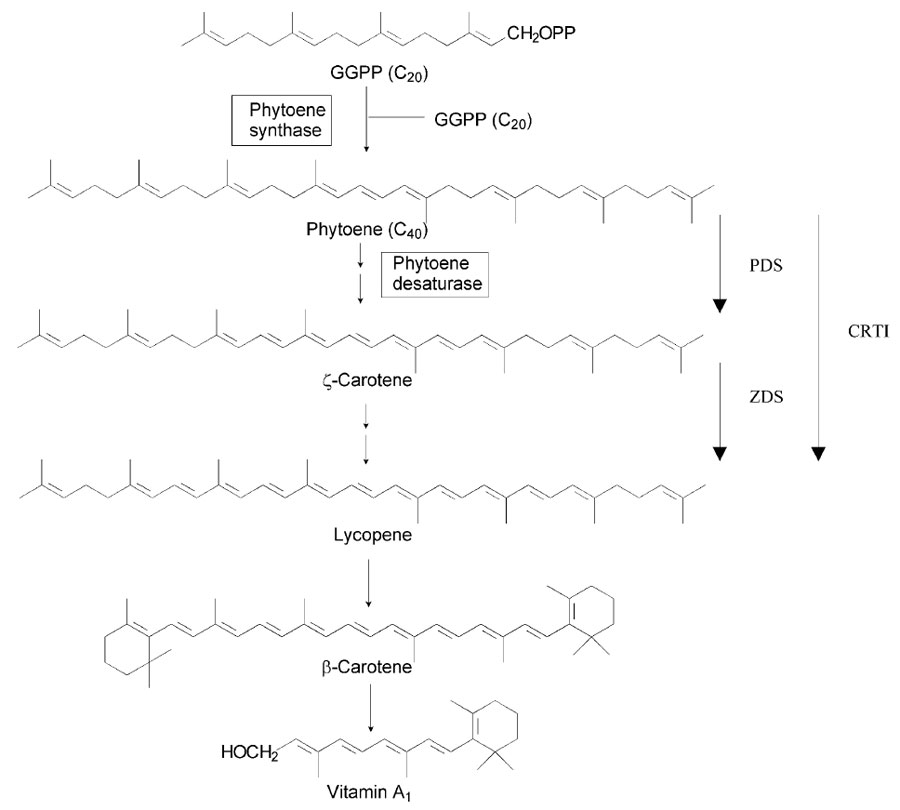
Fig. 6.3 Biosynthetic pathway of vitamin A from geranylgeranyl pyrophosphate (GGPP). Vertical arrows indicate steps catalysed by phytoene desaturase (PDS), carotene desaturase (ZDS), and Erwinia uredovora phytoene desaturase (CRTI).
The edible part of rice grains, the endosperm, lacks an essential nutrient as vitamin A. A diet mostly based on rice consumption may eventually cause vitamin A deficiency. It is estimated that improved vitamin A nutrition could prevent worldwide 1–2 million deaths annually among children. A very promising achievement has been the introduction of genes into rice that enabled the biosynthesis in the endosperm of β-carotene, the precursor of vitamin A (Ye et al., 2000). β-carotene is synthesised from the precursor geranylgeranyl bisphosphate which is converted to the colourless phytoene by the enzyme phytoenesynthase (Fig. 6.3). The phytoene under goes four desaturations to form lycopene, which is red and gives colour to ripened tomato fruits. Further cyclisation of lycopene results in the formation of β-carotene. Immature rice endospermis capable of synthesising geranylgeranyl biphosphate which can be used to produce phytoene by expression of a phytoene synthase gene. The introduction of three genes in rice via Agrobacterium allowed the expression of the entire β-carotene pathway into the endosperm.
Fig. 6.3 Biosynthetic pathway of vitamin A from geranylgeranyl pyrophosphate (GGPP). Vertical arrows indicate steps catalysed by phytoene desaturase (PDS), carotene desaturase (ZDS), and Erwinia uredovora phytoene desaturase (CRTI).
Another lipid-soluble vitamin whose function is linked to an antioxidant role is vitamin E (α-tocopherol). Daily intake of this vitamin in excess of a recommended minimum is associated with decreased incidence of several diseases. Plant oils are the main source of dietary vitamin E and they generally have a high content of the vitamin E precursor γ-tocopherol. Overexpression of γ-tocopherol methyl transferase greatly increased the seed level of α-tocopherol in the model plantArabidopsis thaliana (Shintani and DellaPenna, 1998). This process seems ready to be eventually applied to some commercial crops in the future.
A high risk of iron deficiency has been reported when vegetables are the major components in the diet. Although some plants are rich in this element, its availability is limited by the fact that the same plants contain oxalic acid and phytate-like substances that may complex this element. Some studies have shown that oral administration of ferritin, a protein used by plants and animals to store iron, can provide the iron needed to treat anaemia in rats. With this information, rice has been transformed with a soybean gene encoding ferritin, under the control of a seed-specific promoter (Goto et al. 1999). Transgenic rice plants accumulated ferritin in the endosperm tissue and up to three-fold levels of iron in comparison to normal seeds. Interestingly, plants overexpressing ferritin have been reported to be tolerant to oxidative damage and pathogens. This seems to be an additional agronomic trait for these transgenic plants´k (Dea et al. 1999).
Support our developers




