Dinophyta
The most common type of extrusomes, of almost universal occurrence in the motile phase of these algae, are trichocysts, that is, rod-shaped bodies that, when mature, usually lie in the amphiesma perpendicular to the cell membrane (Figure 2.84). The shaft is a paracrystalline, proteinaceous rod a few micrometers long and rectangular in cross-section. At its distal end it extends as a
group of twisted fibers. The whole is enclosed within a membranous sac, and there is a sheathing material between the rod and the membrane. The tip of the sac is in contact with the cell membrane, passing through the amphiesmal vesicles, and the thecal plates, if present. Thrichocysts are formed in the vicinity of the Golgi body and move to the cell periphery. They are discharged apparently by a rapid hydration process, the discharged structures measuring up to 200 µm in length. Their function is unknown, but it is assumed to be defensive, excretory, or both.
A less ordered type of extrusomes in dinoflagellates is the mucocyst, a simple sac with granular content, associated with the release of mucoid material. They are located just beneath the cell membrane, and are often aggregated in the region of the sulcus; their release in some species is correlated with the psamnophilous existence of these algae, facilitating the attachment of the cells to sand grains along the seashore.More elaborate extrusomes termed nematocysts are found in genera such as
Polykrikos kofoidi and Nematodinium; usually only about eight to ten nematocysts are present per cell. These organelles are larger than trichocysts and can reach 20 µm in length. They are conical, fluid-filled sacs with a capitate blunt end. Most of the body consists of a large posterior chamber, from which a smaller anterior chamber is isolated; the whole structure is capped by a lid-like operculum. A sharp stylet in the anterior chamber is connected to a tubular filament in the posterior chamber (Figure 2.85).
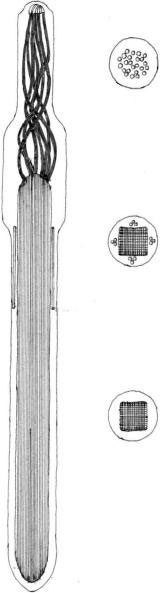
FIGURE 2.84 Undischarged trichocyst of Goniaulax poliedra in longitudinal and transverse sections.
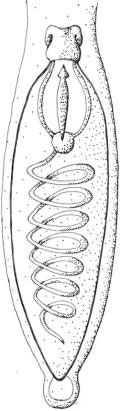
FIGURE 2.85 Nematocysts of Polykrikos kofoidi.
Feeding structures are present in dinoflagellates, depending on the type of feeding mechanisms present: feeding tube (peduncle, or tentacle) and feeding veil (pallium). Tube feeding is commonly found among both naked and thecate species of dinoflagellates (e.g.,
Dinophysis,
Amphidinium,
Gyrodinum, and
Peridinopsis). Cell membrane is extruded from the cell to form a tube, which can engulf whole cells or penetrate prey cell walls and suck in prey cytoplasma.
Pallium feeding has been described only for heterotrophic thecate species such as
Protoperidinium and
Diplopsalis. The prey is captured via a primary attachment filament; an extension of the cytoplasm then emerges from the region of the sulcus-cingulum, which encloses the prey as a veil (pallium). Enzymatical digestion of the prey cytoplasm is brought about inside the veil and the products are then transported to the predator.
Direct engulfment is mainly found among naked species (e.g.,
Gyrodinium,
Gymnodinium, and
Noctiluca); recently, however, some thecate species have been shown to use this feeding mechanism as well. Feeding behavior in dinoflagellates involves several steps prior to actual ingestion, including precapture, capture, and prey manipulation. As feeding mechanisms allow the ingestion of relatively large preys, or parts thereof, dinoflagellates are regarded as raptorial feeders. While prey size plays an important role for the ability of dinoflagellates to ingest food, this cannot alone explain prey preferences actually found. Some dinoflagellate species can be very selective in their choice of prey, while others show a remarkable versatility.




