Outside the Cell
Cell surface forms the border between the external word and the inside of the cell. It serves a
number of basic functions, including species identification, uptake and excretion/secretion of
various compounds, protection against desiccation, pathogens, and predators, cell signaling and
cell–cell interaction. It serves as an osmotic barrier, preventing free flow of material, and as a selective
barrier for the specific transport of molecules. Algae, besides naked membranes more typical of
animal cells and cell walls similar to those of higher plant cells, possess a wide variety of cell surfaces.
The terminology used to describe cell surface structures of algae is sometimes confusing; to
avoid this confusion, or at least to reduce it, we will adopt a terminology mainly based on that of
Presig et al. (1994).
Cell surface structures can be grouped into four different basic types:
- Simple cell membrane (Type 1)
- Cell membrane with additional extracellular material (Type 2)
- Cell membrane with additional intracellular material in vesicles (Type 3)
- Cell membrane with additional intracellular and extracellular material (Type 4)
Type 1: Simple Cell Membrane
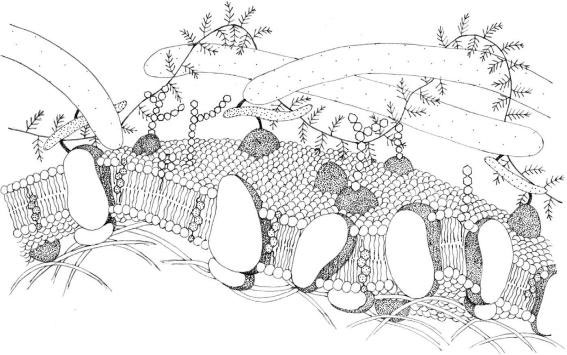
FIGURE 2.1 Schematic drawing of a simple cell membrane.
This cell surface consists of a simple or modified plasma membrane. The unit membrane is a lipid
bilayer, 7–8 nm thick, rich of integral and peripheral proteins. Several domains exist in the membrane,
each distinguished by its own molecular structure. Some domains have characteristic carbohydrate
coat enveloping the unit membrane. The carbohydrate side chains of the membrane
glycolipids and glycoproteins form the carbohydrate coat. Difference in thickness of plasma membrane
may reflect differences in the distribution of phospholipids, glycolipids, and glycoproteins
(Figure 2.1).
FIGURE 2.1 Schematic drawing of a simple cell membrane.
A simple plasma membrane is present in the zoospores and gametes of Chlorophyceae, Xanthophyceae
(Heterokontophyta), and Phaeophyceae (Heterokontophyta), in the zoospores of the
Eustigmatophyceae (Heterokontophyta), and in the spermatozoids of Bacillariophyceae (Heterokontophyta).
This type of cell surface usually characterizes very short-lived stages and, in this
transitory naked phase, the naked condition is usually rapidly lost once zoospores or gametes
have ceased swimming and have become attached to the substrate, as wall formation rapidly
ensues. A simple cell membrane covers the uninucleate cells that form the net-like plasmodium
of the Chlorarachniophyta during all their life history. Most Chrysophyceae occur as naked
cells, whose plasma membrane is in direct contact with water, but in
Ochromonas, the membrane
is covered with both a carbohydrate coat and surface blebs and vesicles, which may serve to trap
bacteria and other particles that are subsequently engulfed as food. The properties of the membrane
or its domains may change from one stage in the life cycle to the next.
Type 2: Cell Surface with Additional Extracellular Material
Extracellular matrices occur in various forms and include mucilage and sheaths, scales, frustule,
cell walls, loricas, and skeleta. The terminology used to describe this membrane-associated material
is quite confusing, and unrelated structures such as the frustule of diatoms, the fused scaled covering
of some prasynophyceae, and the amphiesma of dinoflagellates have been given the same name,
that is, theca. Our attempt has been to organize the matter in a less confusing way (at least in our
opinion).
Mucilages and Sheaths
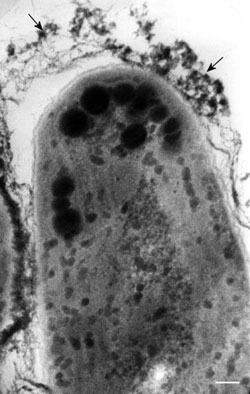
FIGURE 2.2 Transmission electron microscopy image of the apical cell of Leptolyngbya spp. trichome in longitudinal section. The arrows point to the mucilaginous sheath of this cyanobacterium. Inside the cell osmiophylic eyespot globules are present. (Bar: 0.15 µm). (Courtesy of Dr. Patrizia Albertano.)
These are general terms for some sort of outer gelatinous covering present in both prokaryotic and
eukaryotic algae. Mucilages are always present and we can observe a degree of development of
a sheath that is associated with the type of the substrate the cells contact (Figure 2.2). All cyanobacteria secrete a gelatinous material, which, in most species, tends to accumulate around the
cells or trichome in the form of an envelope or sheath. Coccoid species are thus held together to
form colonies; in some filamentous species, the sheath may function in a similar manner, as in
the formation of Nostoc balls, or in development of the firm, gelatinous emispherical domes of
the marine Phormidium crosbyanum. Most commonly, the sheath material in filamentous species
forms a thick coating or tube through which motile trichomes move readily. Sheath production is a
continuous process in cyanobacteria, and variation in this investment may reflect different physiological stages or levels of adaptation to the environment. Under some environmental conditions the sheath may become pigmented, although it is ordinarily colorless and transparent. Ferric hydroxide
or other iron or
metallic salts may accumulate in the sheath, as well as pigments originating within
the cell. Only a few cyanobacterial exopolysaccharides have been defined structurally; the sheath of
Nostoc commune contains cellulose-like glucan fibrils cross-linked with minor monosaccharides,
and that of Mycrocystis flos-aquae consists mainly of galacturonic acid, with a composition
similar to that of pectin. Cyanobacterial sheaths appear as a major component of soil crusts
found throughout the world, from hot desert to polar regions, protecting soil from erosion,
favoring water retention and nutrient bio-mobilization, and affecting chemical weathering of the
environment they colonize.
FIGURE 2.2 Transmission electron microscopy image of the apical cell of Leptolyngbya spp. trichome in longitudinal section. The arrows point to the mucilaginous sheath of this cyanobacterium. Inside the cell osmiophylic eyespot globules are present. (Bar: 0.15 µm). (Courtesy of Dr. Patrizia Albertano.)
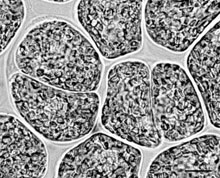
FIGURE 2.3 Palmelloid phase of Euglena gracilis. (Bar: 10 µm.)
In eukaryotic algae, mucilages and sheaths are present in diverse divisions. The most common
occurence of this extracellular material is in the algae palmelloid phases, in which non-motile cells
are embedded in a thick, more or less stratified sheath of mucilage. This phase is so-called because
it occurs in the genus Palmella (Chlorophyceae), but it occurs also in other members of the same
class, such as Asterococcus sp., Hormotila sp., Spirogyra sp., and Gleocystis sp. A palmelloid phase
is present also in Chroomonas sp. (Cryptophyceae) and in Gleodinium montanum vegetative cells
(Dynophyceae) and in Euglena gracilis (Euglenophyceae) (Figure 2.3).
Less common are the cases
in which filaments are covered by continuous tubular layers of mucilages and sheath. It occurs in
the filaments of Geminella sp. (Chlorophyceae). A more specific covering exists in the filaments of
Phaeothamnion sp. (Chrysophyceae), because under certain growth conditions, cells of the filaments
dissociate and produce a thick mucilage that surround them in a sort of colony resembling
the palmelloid phase.
FIGURE 2.3 Palmelloid phase of Euglena gracilis. (Bar: 10 µm.)




