The Circulatory Organs
The heart of the vertebrate embryo is at first a simple tube, the anterior end of which passes into a cardiac aortic trunk, while the posterior end is continuous with the great veins which bring back blood from the umbilical vesicle - the omphalomeseraic veins.The cardiac aorta immediately divides into two branches, each of which ascends, in the first visceral arch, in the form of a forwardly convex aortic arch, to the under side of the rudimentary spinal column, and then runs, parallel with its fellow, to the hinder part of the body, as a primitive subvertebral aorta. The two primitive aortse very soon coalesce throughout the greater part of their length into one trunk, the definitive subvertebral aorta; but the aortic arches, separated by the alimentary tract, remain distinct. Additional arterial trunks, to the number of four in the higher Vertebrata, and more in the lower, are successively developed, behind the first, in the other visceral arches, and further connect the cardiac and subvertebral aortae.
In the permanently branchiate Vertebrata, the majority of these aortic arches persist, giving off vessels to the branchial tufts, and becoming converted into afferent and efferent trunks, which carry the blood to and take it from these tufts. (Fig. 25, A, B, C, D, E.)
In the higher Amphibia, which, though branchiate in the young state, become entirely air-breathers in the adult condition, such as the Batrachia (Fig. 35, F) and Caecilia, the permeable aortic arches are reduced to two (the middle pair of the three which supply the external gills, and the fourth pair of embryonic aortic arches) by the obliteration of the cavities of the dorsal ends of the others. Of the posterior arches, the remains of the fifth and sixth become the trunks which give off the pulmonary arteries, and, in the Batrachia, cutaneous branches. The anterior, or third, primitive aortic arch becomes the common carotid trunk, and ends in the carotid gland, whence the internal and external carotids arise. In those Vertebrata which never possess gills, the arches become reduced either to two pair, as in some Lacertilia; or to one pair, as in other Reptilia; or to a single arch, as in Aves and Mammalia. The aortic arches thus retained are, in the Lizards in question, the third and the fourth pairs in order from before backward; but the fourth pair only, in other Reptiles; in Birds, the right arch only of the fourth pair; and in Mammals, the left arch only of the fourth pair. The fifth pair of arches give off the pulmonary arteries, the so-called "ductus arteriosus" representing the remains of the primitive connection of these arches with the fourth pair and the subvertebral aorta. The dorsal ends of the first, second, and third arches become obliterated; but their cardiac ends, and the branches which they give off, become the arteries of the head and upper extremities.
The embryonic aorta gives off omphalomeseraic branches (Fig. 26, o) to the umbilical vesicle; and ends, at first, in the hypogastric arteries (which are distributed to the allantois in the abranchiate Vertebrata), and a median caudal continuation.
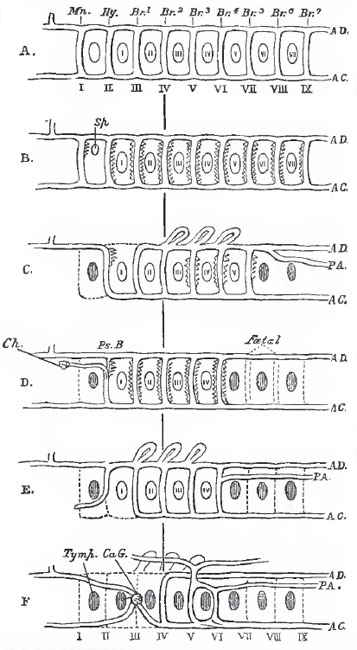 |
Fig. 25. - A diagram Intended to show the manner in which the aortic
arches become modified in the series of the Vertebrata.
|
The blood of the allantois is returned by the umbilical vein, or veins (Fig. 26, u), which are formed in the anterior wall of the abdomen, and open into the venous sinus before mentioned. The blood of the posterior extremities and kidneys is, after a while, brought to the same point by a special median vein, the vena cava inferior (Fig. 26, cv).
The development of the liver effects the first great change in the arrangements now described. It, as it were, interrupts the course of the omphalomeseraic vein, which is not only the vein of the umbilical sac but also that of the intestine, and converts it into a meshwork of canals, which communicate, on one side, with the cardiac part of the vein, and, on the other side, with its intestinal part. The latter is thus converted into the vena portae (Fig. 26, vp), distributing the blood of the stomaoh and intestines to the liver; while the former becomes the hepatic vein (vh), carrying the hepatic blood to the inferior cava, and thence to the heart.
The umbilical vein further gives a branch to the liver; while, on the other hand, it communicates directly with the venous sinus (now almost merged in the vena cava inferior) by a trunk called ductus venosus (Fig. 26, Dv).
When the umbilical vesicle and allantois cease to have any further import, as at birth, or before, the omphalomeseraic arteries have become intestinal arteries, and the omphalomeseraic vein, the vena portae. The hypogastric arteries are obliterated, except so much of them as is converted into the common iliac arteries. The umbilical vein, or veins, also disappear, or are represented by mere ligaments.
Of the three veins which open into the venous sac - viz, the inferior cava, and the right and left ductus Cuvieri - all may persist, the latter receiving the title of right and left supperior cavae. Or, as very often happens in the higher Vertebrata, the left ductus Cuvieri becomes more or less obliterated; the veins which properly open into it acquiring a connection with the right ductus, which then remains as the sole superior cava. The posterior cardinal veins give off anastomosing branches, which are converted into the vence azygos; the anterior cardinal veins become metamorphosed into the external jugular veins and vence innominatoe.
In Fishes, the sinus venosus and the cardinal veins persist throughout life; but the anterior cardinal veins, which bring back the blood from the head and from the anterior extremities, are called vence jugulares.
The caudal veins are either directly continued into the cardinal veins, as in Marsipobranchii and Elasmohranchii, or branch out into the kidneys, as in many Teleostei. In either case the efferent renal veins open into the cardinal veins.
The portal veins, conveying the blood of the chylopoietic viscera, and sometimes that of other organs and of the abdominal walls, may be one or many. In Amphioxus and Myxine the vein is rhythmically contractile, and forms a portal heart.
In most Amphibia and Reptilia the sinus venosus persists, and is rhythmically contractile, valves being placed at its opening into the right auricle.
The anterior cardinal veins are represented by jugular veins, the posterior cardinal by vertebral veins; these, and the veins of the anterior extremities, when they are present, pour their blood into the ductus Cuvieri, which are now termed anterior vence cavce.
The vena cava inferior takes its origin chiefly by the coalescence of the efferent veins of the kidneys and reproductive organs, and does not always receive the whole of the hepatic veins - more or fewer of the latter opening independently into the sinus venosus.
The blood which leaves the kidneys by its efferent veins is supplied, not only by the renal arteries, but by the veins of the caudal region, and of the hinder extremities, which branch out like a vena portae in the substance of the kidneys. This renal portal system is less developed in Reptilia than in Amphibia. All the blood of the posterior extremities and caudal region does not traverse the kidneys, however, more or less of it being led away by great branches of the iliac veins, which run along the anterior wall of the abdominal cavity, either as two trunks, or united into one. These vence abdominales anteriores are eventually distributed to the liver, along with the branches of the proper vence portoe.
 |
| Fig. 26. - Diagram of the arrangement of the principal vessels in a human foetus. - II, the heart; TA, the eortic trunk or cardiac aorta; c, the common carotid; c', the external carotid; c", the internal carotid; s, subclavian; v, vertebral artery; 1, 2, 3, 4, 5, the aortic arches - the persistent left aortic arch is hidden. A', subvertebral aorta; o, omphalomeseraic artery, going to the umbilical vesicle v, with its vitelline duct dv; o, omphalomeseraic vein; vp, the vena portae; L, the liver; uu, tho hypogastric or umbilical arteries, with their placental ramifications, u", u",u' the umbilical vein; Dv, the ductus venosus; vh, tbe hepatic vein; cv, the vena cava inferior; vil, the iliac veins; az, a vena azygos; vc', a vena cardinalis posterior; DC, a ductus Cuvieri; the anterior cardinal vein is seen commencing in the head and running down to the ductus Onvieri on the under side of the numbers 1, 2, 3, 4, 5 ; P, the lungs. |
The caudal and pelvic veins unite into three principal trunks, of which one is median and two are lateral. The median enters into the portal system. The lateral branches pass along and through the kidney, receiving veins from it, but giving none to it; and eventually, after receiving the ischiatic veins, unite with the crural veins to form the common iliacs. Thus there is no renal portal system in birds.
In Mammalia, the sinus venosus is not distinct from the right article. The anterior cavce are frequently reduced to one, the right. The vena cava inferior commences in the caudal region, and receives all the blood of the posterior moiety of the body, except so much as is carried away by the azygous veins. The anterior abdominal veins are represented only during foetal life, by the umbilical vein or veins. The efferent veins of the kidneys open directly into the trunk of the inferior vena cava, and the portal vein is composed exclusively of radicles proceeding from the chylopoietic viscera.
Many of the veins of Amphioxus, the portal vein of Myxine, dilatations of the caudal vein in the Eel, the venae cavse and the iliac and axillary veins of many Amphibia, the veins of the wing of Bats, possess a rhythmical contractility, which, in combination with the disposition of their valves, assists the circulation of the blood.
In Vertebrata of all classes, and in very diverse parts of the body, both veins and arteries occasionally break up into numerous branches of nearly equal size, which may or may not unite again into larger trunks. These are called retia mirabilia.
Support our developers




