Cytokines
Cytokines
The 1980s saw rapid advances in our knowledge of how cells of immunity communicate with each other. They do this by means of protein hormones called cytokines (Table 37-1). Cytokines can produce their effects on the same cells that produce them, on cells nearby, or on cells distant in the body from those that produced the cytokine.
Interleukins were originally so called because they are synthesized by leukocytes and have their effect on leukocytes. We now know that some other kinds of cells can produce interleukins, and interleukins produced by leukocytes can affect other kinds of cells.
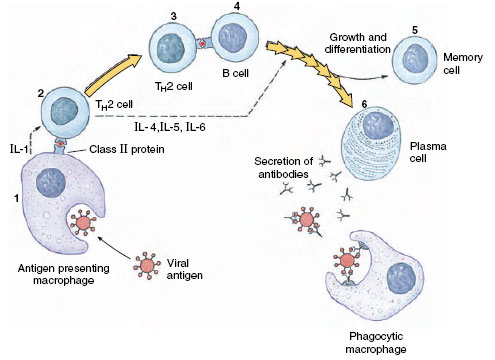
Generation of a Humoral Response: TH2 Arm When an antigen is introduced into a body, it binds to a specific antibody on the surface of an appropriate B cell, but this binding is usually not sufficient to activate the B cell to multiply. Some of the antigen is taken up by antigenpresenting cells (APCs), such as macrophages, that partially digest the antigen. The APCs then incorporate portions of the antigen into their own cell surface, bound in the cleft of MHC II protein (Figures 37-4 and 37-5). That portion of the antigen presented on the surface of the macrophage or other APC is called the epitope (or determinant). The macrophages also secrete IL-1, which stimulates TH2 cells. The specific T-cell receptor for that particular epitope recognizes the epitope bound to the MHC II protein. Binding of the T-cell receptor to the epitope-MHC II complex is enhanced by the coreceptor CD4, which itself binds to the constant portion of the MHC II protein (Figure 37-5). Bound CD4 molecule also transmits a stimulation signal to the interior of the T cell. Activation of the T-cell further requires interaction of additional costimulatory and adhesion signals from other proteins on the surface of the macrophage and T cell. CD8 coreceptors function in a similar way on CD8+ cells; they enhance binding of the T-cell receptor and transmit a stimulatory signal into the T cell.
Many aspects of immunology have been greatly assisted by the discovery of a method for producing stable clones of cells that will produce only one kind of antibody. Such monoclonal antibodies will bind only to one kind of antigenic determinant (most proteins bear many different antigenic determinants and thus stimulate the body to produce complex mixtures of antibodies). Monoclonal antibodies are made by fusing normal antibodyproducing plasma cells with a continuously growing plasma cell line, producing a hybrid of the normal cell with one that can divide indefinitely in culture. This cell line is called a hybridoma. Clones are selected from among the hybrids and are grown to become “factories” that produce almost unlimited quantities of one specific antibody. Hybridoma techniques discovered in 1975 have become one of the most important research tools for the immunologist.
Activated TH2 cells secrete IL-4, IL-5 and IL-6, which activate the B cell that has the same epitope and class II MHC protein on its surface. The B cell multiples rapidly and produces many plasma cells, which secrete large quantities of antibody for a period of time, then die. Thus if we measure the concentration of antibody (titer) soon after the antigen is injected, we can detect little or none. Titer then rises rapidly as plasma cells secrete the antibody, and it may decrease somewhat as they die and antibody is degraded (Figure 37-6). However, if we give another dose of antigen (challenge), there is little or no lag, and antibody titer rises quicker to a higher level than after the first dose. This is the secondary or anamnestic response, and it occurs because some of the activated B cells gave rise to long-lived memory cells. There are many more memory cells present in the body than the original B lymphocyte with the appropriate antibody on its surface, and they rapidly multiply to produce additional plasma cells. Existence of the anamnestic response has great practical value because it is the basis for protective vaccines.
Cell-Mediated Response: TH1 Arm Many immune responses involve little, if any, antibody and depend on the action of cells only. In cell-mediated immunity (CMI) the epitope of an antigen is also presented by ACPs, but the TH1 arm of the immune response is activated and the TH2 arm suppressed. Effector cells are macrophages, PMNs, cytotoxic T cells, and activated natural killer cells. The specific interaction of lymphocyte and antigen that generates a CMI greatly influences subsequent events in the nonspecific response we call inflammation.
Only a few years ago transplantation of organs from one person to another seemed impossible. Then physicians began to transplant kidneys and depress the immune response to the recipient. It was very difficult to immunosuppress the recipient enough that the new organ would not be rejected and at the same time not leave the patient defenseless against infection. Since discovery of a fungus-derived drug called cyclosporine, not only kidneys, but also hearts, lungs, and livers can be transplanted. Cyclosporine inhibits IL-2 and affects CTLs more than THs. It has no effect on other white cells or on healing mechanisms, so that a patient can still mount an immune response but not reject the transplant. However, the patient must continue to take cyclosporine because if the drug is stopped, the body will recognize the transplanted organ as foreign and reject it.
Like humoral immunity, CMI shows a secondary response due to large numbers of memory T cells produced from the original activation. For example, a second tissue graft (challenge) between the same donor and host will be rejected much more quickly than the first.
The 1980s saw rapid advances in our knowledge of how cells of immunity communicate with each other. They do this by means of protein hormones called cytokines (Table 37-1). Cytokines can produce their effects on the same cells that produce them, on cells nearby, or on cells distant in the body from those that produced the cytokine.
 |
Interleukins were originally so called because they are synthesized by leukocytes and have their effect on leukocytes. We now know that some other kinds of cells can produce interleukins, and interleukins produced by leukocytes can affect other kinds of cells.
 |
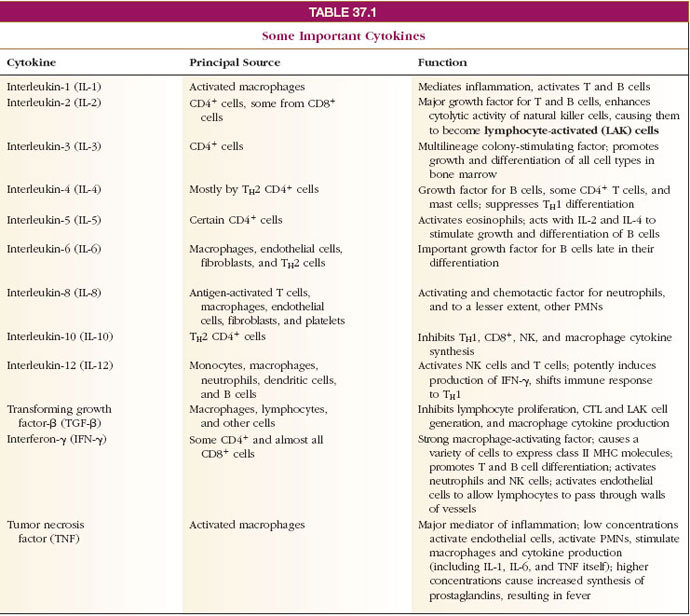
| Figure 37-4 Humoral immune response. (1) Macrophage consumes antigen, partially digests it, and displays portions on its surface, along with class II MHC protein, and secretes interleukin-1 (IL-1). (2) TH2 cell, stimulated by IL-1, recognizes antigen and class II protein on macrophage, is activated, and secretes interleukins 4, 5, and 6 (IL-4, IL-5, IL-6). (3) TH2 then activates B cell which carries antigen and class II protein on its surface. IL-4, IL-5, and IL-6 stimulate proliferation of B cell line. (4) Activated B cells finally produced many plasma cells that secrete antibody. (5) Some B-cell progeny become memory cells. (6) Antibody produced by plasma cells binds to antigen and stimulates macrophages to consume antigen (opsonization). |
Generation of a Humoral Response: TH2 Arm When an antigen is introduced into a body, it binds to a specific antibody on the surface of an appropriate B cell, but this binding is usually not sufficient to activate the B cell to multiply. Some of the antigen is taken up by antigenpresenting cells (APCs), such as macrophages, that partially digest the antigen. The APCs then incorporate portions of the antigen into their own cell surface, bound in the cleft of MHC II protein (Figures 37-4 and 37-5). That portion of the antigen presented on the surface of the macrophage or other APC is called the epitope (or determinant). The macrophages also secrete IL-1, which stimulates TH2 cells. The specific T-cell receptor for that particular epitope recognizes the epitope bound to the MHC II protein. Binding of the T-cell receptor to the epitope-MHC II complex is enhanced by the coreceptor CD4, which itself binds to the constant portion of the MHC II protein (Figure 37-5). Bound CD4 molecule also transmits a stimulation signal to the interior of the T cell. Activation of the T-cell further requires interaction of additional costimulatory and adhesion signals from other proteins on the surface of the macrophage and T cell. CD8 coreceptors function in a similar way on CD8+ cells; they enhance binding of the T-cell receptor and transmit a stimulatory signal into the T cell.
 |
| Figure 37-5 Interacting molecules during activation of a T helper cell. |
Many aspects of immunology have been greatly assisted by the discovery of a method for producing stable clones of cells that will produce only one kind of antibody. Such monoclonal antibodies will bind only to one kind of antigenic determinant (most proteins bear many different antigenic determinants and thus stimulate the body to produce complex mixtures of antibodies). Monoclonal antibodies are made by fusing normal antibodyproducing plasma cells with a continuously growing plasma cell line, producing a hybrid of the normal cell with one that can divide indefinitely in culture. This cell line is called a hybridoma. Clones are selected from among the hybrids and are grown to become “factories” that produce almost unlimited quantities of one specific antibody. Hybridoma techniques discovered in 1975 have become one of the most important research tools for the immunologist.
 |
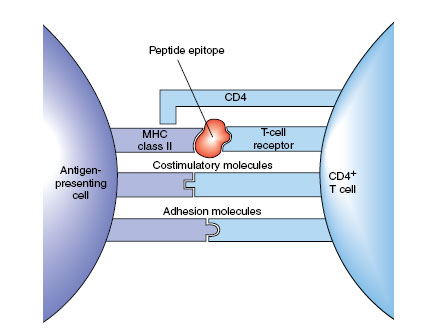
| Figure 37-6 Typical immunoglobulin response after primary and challenge immunizations. The secondary response is a result of the large numbers of memory cells produced after the primary B-cell activation. |
Activated TH2 cells secrete IL-4, IL-5 and IL-6, which activate the B cell that has the same epitope and class II MHC protein on its surface. The B cell multiples rapidly and produces many plasma cells, which secrete large quantities of antibody for a period of time, then die. Thus if we measure the concentration of antibody (titer) soon after the antigen is injected, we can detect little or none. Titer then rises rapidly as plasma cells secrete the antibody, and it may decrease somewhat as they die and antibody is degraded (Figure 37-6). However, if we give another dose of antigen (challenge), there is little or no lag, and antibody titer rises quicker to a higher level than after the first dose. This is the secondary or anamnestic response, and it occurs because some of the activated B cells gave rise to long-lived memory cells. There are many more memory cells present in the body than the original B lymphocyte with the appropriate antibody on its surface, and they rapidly multiply to produce additional plasma cells. Existence of the anamnestic response has great practical value because it is the basis for protective vaccines.
Cell-Mediated Response: TH1 Arm Many immune responses involve little, if any, antibody and depend on the action of cells only. In cell-mediated immunity (CMI) the epitope of an antigen is also presented by ACPs, but the TH1 arm of the immune response is activated and the TH2 arm suppressed. Effector cells are macrophages, PMNs, cytotoxic T cells, and activated natural killer cells. The specific interaction of lymphocyte and antigen that generates a CMI greatly influences subsequent events in the nonspecific response we call inflammation.
Only a few years ago transplantation of organs from one person to another seemed impossible. Then physicians began to transplant kidneys and depress the immune response to the recipient. It was very difficult to immunosuppress the recipient enough that the new organ would not be rejected and at the same time not leave the patient defenseless against infection. Since discovery of a fungus-derived drug called cyclosporine, not only kidneys, but also hearts, lungs, and livers can be transplanted. Cyclosporine inhibits IL-2 and affects CTLs more than THs. It has no effect on other white cells or on healing mechanisms, so that a patient can still mount an immune response but not reject the transplant. However, the patient must continue to take cyclosporine because if the drug is stopped, the body will recognize the transplanted organ as foreign and reject it.
Like humoral immunity, CMI shows a secondary response due to large numbers of memory T cells produced from the original activation. For example, a second tissue graft (challenge) between the same donor and host will be rejected much more quickly than the first.
Support our developers




