Immunity in Invertebrates
Immunity in
Invertebrates
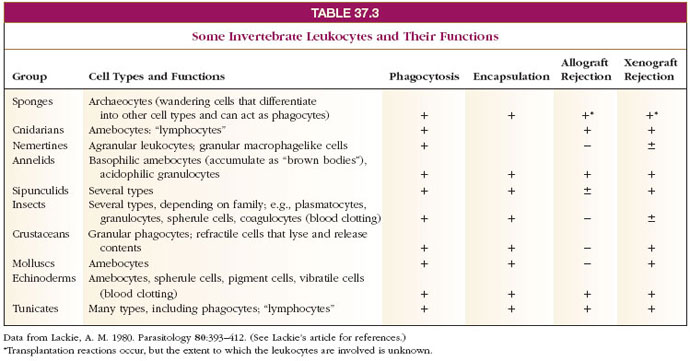
One of the principal tests of the ability of invertebrate tissues to recognize nonself is by grafting of a piece of tissue from another individual of the same species (allograft) or a different species (xenograft) onto the host. If the graft grows in place with no host response, the host tissue is treating it as self, but if cell response and rejection of the graft occur, the host exhibits immune recognition. Most invertebrates tested in this way reject xenografts; and almost all can reject allografts to some degree (Table 37-3). Interestingly, nemertines and molluscs apparently do not reject allografts. It is curious that some animals with quite simple body organization, such as Porifera and Cnidaria, can reject allografts; this response may be an adaptation to avoid loss of integrity of the individual sponge or colony under conditions of crowding, with attendant danger of overgrowth or fusion with other individuals. American cockroaches (Periplaneta americana) reject allografts from the same source more quickly upon second exposure; thus they show at least short-term immunological memory.
Hemocytes of molluscs release degradative
enzymes during phagocytosis
and encapsulation, and bactericidal
substances occur in body fluids of a
variety of invertebrates. Substances
functioning as opsonins have been
reported in annelids, insects, crustaceans,
echinoderms, and molluscs.
Bacterial infection in some insects
stimulates production of antibacterial
proteins, but these proteins show broadspectrum
activity and are not specific
for a single infective agent. Specific,
induced responses that demonstrate
memory upon challenge, resembling
acquired immunity of vertebrates, have
been found in American cockroaches.
Contact with infectious organisms can bring the defense systems of snails into enhanced levels of readiness that last for up to two months or more. Susceptibility of snail hosts of the trematode Schistosoma mansoni depends heavily on genotype of the snail. Excretory/secretory products of the trematode stimulate motility of hemocytes from resistant snails but inhibit motility of hemocytes from susceptible hosts. Hemocytes from resistant snails encapsulate the trematode larva and apparently kill it with superoxide and H2O2 and then destroy it by phagocytosis (Figure 37-9). It appears that the cytokine interleukin-1 is present in resistant snails and is responsible for activating the hemocytes.
One of the principal tests of the ability of invertebrate tissues to recognize nonself is by grafting of a piece of tissue from another individual of the same species (allograft) or a different species (xenograft) onto the host. If the graft grows in place with no host response, the host tissue is treating it as self, but if cell response and rejection of the graft occur, the host exhibits immune recognition. Most invertebrates tested in this way reject xenografts; and almost all can reject allografts to some degree (Table 37-3). Interestingly, nemertines and molluscs apparently do not reject allografts. It is curious that some animals with quite simple body organization, such as Porifera and Cnidaria, can reject allografts; this response may be an adaptation to avoid loss of integrity of the individual sponge or colony under conditions of crowding, with attendant danger of overgrowth or fusion with other individuals. American cockroaches (Periplaneta americana) reject allografts from the same source more quickly upon second exposure; thus they show at least short-term immunological memory.
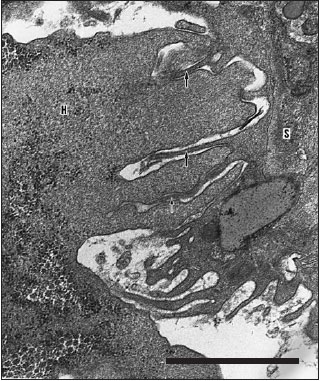 |
| Figure 37-9 Electron micrograph of a
hemocyte (H) from a schistosome-resistant strain of a snail attacking a a Schistosoma mansoni larva (S) under in vitro conditions. Note the hemocyte processes apparently engaged in phagocytosis of portions of the larval tegument (arrows). (Scale bar = 1 µm.) |
Contact with infectious organisms can bring the defense systems of snails into enhanced levels of readiness that last for up to two months or more. Susceptibility of snail hosts of the trematode Schistosoma mansoni depends heavily on genotype of the snail. Excretory/secretory products of the trematode stimulate motility of hemocytes from resistant snails but inhibit motility of hemocytes from susceptible hosts. Hemocytes from resistant snails encapsulate the trematode larva and apparently kill it with superoxide and H2O2 and then destroy it by phagocytosis (Figure 37-9). It appears that the cytokine interleukin-1 is present in resistant snails and is responsible for activating the hemocytes.
Support our developers




