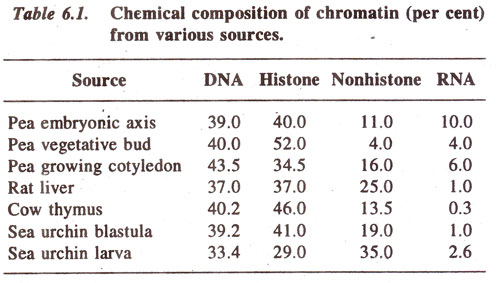
The major chemical components of chromosomes are DNA, RNA, histone proteins and nonhistone proteins. Calcium is also present in addition to these constituents. The relative proportions of different components are given in Table 6.1 for a variety of organisms, which suggest that there is more protein than DNA in chromatin in all cases.
DNA. As we know, DNA is the most important of chemical components of chromatin, since it plays the central role of controlling heredity. Quantitative measurements of DNA have been made in a large number of cases which are reviewed by
H. Rees and
R.N. Jones in 1972 (Intern. Rev. Cytol.). The most convenient measurement of DNA is picogram (10
-12 g) which is equivalent to 31 cm of double helical DNA (for details of the double helical structure of DNA, consult
Chemistry of the Gene 1. Nucleic Acids and Their Structure). It has been found that quantity of DNA varies greatly in cells from different kinds of organisms. The haploid genome of mammals usually contains 1000 times DNA content of bacteria. Other eukaryotes may similarly have 10 to 100 times the bacterial DNA content.
It is interesting to note that a human diploid cell has 174 cm (5-6 picograms) of DNA, so that all cells in a human being may have DNA equal to 2.5 x 10
10 km (100 g), a length which is equal to 100 times the distance from earth to sun. Similarly a diploid cell of
Trillium has 37 meters (120 picograms) and that of
Drosophila salivary glands has 91 meters (293 picograms) of DNA. In comparison of these enormous lengths, the DNA of bacteria measures only 1.1 mm-1.4 mm.
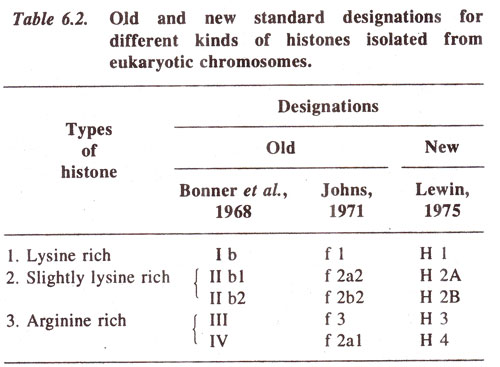
Histones. There are five fractions of histones, which have been differently designated (Table 6.2).
H1 histone is most easily removed and so is least tightly bound. This may thus be concerned with holding together a chromosome fibre. H3 and H4 are extremely conserved, having same structure in different species and should thus have a common structural role.
Histones, isolated from diverse materials showed considerable similarity. It is also assumed that general similarities in histones have been conserved during evolution. This feature alone suggested that these proteins should play a structural role rather than a regulatory role. However, some important chromatin reconstitution and other experiments conducted in 1960s and 1970s established that histones do play a regulatory role. This regulatory role of histones is more of general nature rather than specific and is exercised by repressing the activity of genes.
Non-histones. The non-histone proteins display more
but still limited diversity. In a variety
of organisms, number of non-histones can vary from 12 to little more than 20. Heterogeneity of these proteins suggested that these proteins are not as conserved in evolution as histones. These non-histone proteins differ even between different tissues of the same organism suggesting that they regulate the activity of specific genes. Chromatin constitution experiments described in 1973 by
R.S. Gilmour and
J. Paul of Institute for Cancer Research at Glasgow (U.K.), established that specific non-histone proteins switch on specific genes. The results of these experiments were later confirmed in a number of cases
(Barrett et al., 1974;
Groner et al., 1975).