The gene frequencies may change due to selection in favour of one of the two alleles of a gene. For instance, if individuals with allele
'A' are more successful in reproduction than the individuals with
'a', the frequency of the former will be higher. There may be a number of mechanisms which may influence the reproductive success of a genotype and all of them collectively will determine the degree of selection advantage or disadvantage. The selection can be artificial or natural depending upon whether it is being controlled by human beings or by nature respectively. The factors influencing the selection may include temperature, humidity fluctuations, availability of food, difference is sexual attractions, etc. This selection can be illustrated with the help of an experiment conducted by
Harlap and
Martini in 1938, the results of which are presented in Table 46.4. They used ten distinguishable varieties of barley and mixed equal quantities of seed from each of them. The mixture was grown at five different locations in U.S.A.
The experiment was continued for 4-12 years by continuous harvesting and resowing. If there were no selection against or in favour of any variety, one would expect that even after 4-12
years, the proportion of all ten varieties should be equal at each location, Contrary to this, when at each location, 500 plants taken at random were classified into the ten varieties, it was found that different varieties predominated at different locations as shown in Table 46.4. This is a clear evidence of selection having operated in different locations in favour of different varieties. In view of this, the selection can be regarded as a process of scrutiny which rewards vigorous varieties with reproductive success. There are several aspects of selection, like
fitness, gametic selection and
zygotic selection, which need separate discussions.
From data of Harlan and Martini, 1938.
* The initial population in each locality was founded from the same mixture of seeds of all varieties.
Fitness. When one genotype can produce more offspring than the other in the same environment, it means relative reproductive success, which is called
'fitness or
'adaptive value' or
'selective value'. For instance, if a genotype carrying
'A' produces 100 individuals reaching maturity and another genotype carrying
'a' produces 90 individuals in the same environment, this would mean that the reproductive success of
'a' is reduced by 10% or by a fraction of 0.1. This adaptive value, which is also designated as
'w', ranges from 1.0 for most reproductive genotype to zero for lethals. This may also be expressed in the form of selection coefficient or
's' which is defined as a force acting on a genotype to reduce its adaptive value. If the adaptive value
'w' is 1, selection coefficient V would be zero or
vice versa. Algebraically, this can be expressed as
w=l-
s or
s = l-
w. Sometimes, in literature it may be seen that the adaptive value
'w' may exceed 1. It will happen only when a particular individual is assigned a value of
w = 1, rather arbitrarily, so that if another individual is better, the value of
'w' will exceed 1 in such a case.
Gametic selection. If there is no selection among the gametes, one may assume that the gametes carrying dominant and recessive alleles would be equally efficient. However, it is not always necessary that the two types of gametes will be equally efficient. For instance, in a population, let the frequency of
'A' be
'p' and that of
'a' be
'q'. In such a case, if V is the selection coefficient reducing the fitness of deleterious allele
'a', then
wA = I, wa = I - s. After one generation of selection, the following changes can be noticed :
If
'a' is lethal i.e.
s = 1,
q1 = q (1 -
s)= 0. In such a case
'a' will be eliminated in a single generation. But if
's' is less than one,
'a' will persist and its frequency will be reduced by an amount
sq. The frequencies after selection, therefore, will be :
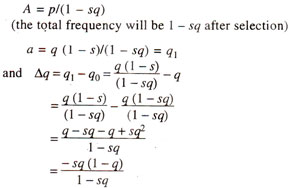
'sq' in the denominator can be ignored, if 5 is small (selection is slow), so that,
Δq = -sq (1 -q)
For
'n' generations this relationship can be summed by methods of
calculus to give.
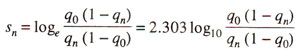
where
q0is the initial frequency of gene and
qn is the frequency of the gene after
n generations of selection.
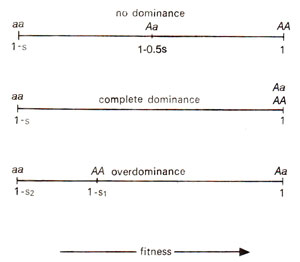
Fig. 46.4. Phenotypic expression of three genotypes AA, Aa, and aa under conditions of no dominance, complete dominance and overdominance (s = selection coefficient).
Zygotic selection. While in gametic selection, the degree of dominance is not important, in case of zygotic selection, degree of dominance also becomes important. Zygote will have three possible genotypes, when a single gene is taken into consideration. These genotypes, as we know, are
AA, Aa and
aa. The expression of no dominance, complete dominance and overdominance leads to three situations illustrated in Figure 46.4, where selection coefficient is expressed as
's'.
Fig. 46.4. Phenotypic expression of three genotypes AA, Aa, and aa under conditions of no dominance, complete dominance and overdominance (s = selection coefficient).
When there is no dominance, if selection coefficient against
'aa' is
's' the fitness of different genotypes will be 1 (
AA)
, 1-0.
5s (
Aa)and 1-s (
aa)
, whereas in case of overdominance it will be expressed as 1 (
Aa)
, 1
-s1(
AA)and 1
-s2(
aa)where
s1 and
s2are the selection coefficients against homogygous dominant and homogygous recessive respectively. The different levels of fitness for the three genotypes can be utilized for calculating the change of frequency of gene 'a', i.e.
'q'. The details of these changes in frequency of
'a' can be available in an advanced text book on quantitative genetics (Falconer's
"Introduction to Quantitative Genetics").






