Heteropolysaccharides
The diversity of heteropolysaccharides is enormous. Distributed throughout the animal and plant kingdom, this class of macromolecules rivals proteins in diversity. Initially thought to function only as structural components, it is now realized that specific sequences may have information content and can serve as recognition or signaling elements. In addition, modifications of saccharides already incorporated into a polymeric chain (sulfation, for example) add an additional level of complexity. The ability of these materials to function as other than strictly physical components necessitates, in many cases, that the saccharide chain have a defined three-dimensional conformation. Much work has been done to model the three-dimensional structureof proteins andmanyproteinstructures havebeen determined by X-ray crystallographic analysis. Much less has been done with complex saccharides in terms of either molecular dynamics or conformational analysis of larger structures. The following discussion is intended to be representative only.In addition to cellulose and xylan, plant cell walls contain a variety of complex heteropolysaccharides including other glucans. Detailed structures have only been determined for a limited number of these structures and their interactions, possibly covalent, with other plant components have rarely been determined in detail. Commercially important are pectins, polygalcturonides, which are key components of jellies and related uronides (gums) used as thickening agents in a broad variety of applications ranging from ice cream manufacture to the production of printing ink. Agars are galactans that are sulfated and also contain 3,6-anhydro-L-galactose, which have gelation properties. They have broad application in microbiology.
Bacteria likewise produce a wide variety of complex, celL-surface polysaccharides. Strains of Streptococcus pneumoniae, the causative agent of bacterial pneumonia, are characterized by a capsular polysaccharide—more than 100 different types are known. Since the capsular material is immunogenic and protective, polyvalent vaccines have been developed that utilize the capsular polysaccharides from common strains. Similarly, organisms responsible for bacterial meningitis have a characteristic polysaccharide capsule that is also immunogenic and protective. Conversely, several bacteria have, as an exterior capsule, saccharides sufficiently similar to those produced in their animal host so as to serve as a mechanism for avoidance of host immune responses. Many gram-positive bacteria have, as essential cellwall components, a complex saccharide structure that contains muramic acid (a condensation product of N-acetylglucosamine and pyruvate) and other amino sugars and is cross-linked by a peptide (Fig. 23).
A widely distributed heteropolysaccharide, hyaluronic acid, has both commerical and biological importance. This molecule, a repeating structure of D-glucuronic acid and N-acetylglucosamine with β-1-3 and β-1-4 linkages, is found in bacterial and animal sources, and it is one of the few complex saccharides not covalently linked to protein (Fig. 24). Molecular weights vary depending on source but often exceed two million. The polyanionic nature of the molecule leads to a relatively extended solution conformation. This coupled with the highly hydrophilic chemistry results in solutions with very high viscosity, an important physical property. This is utilized in treatment of osteoathritis of the knee (an injectable) and in eye surgery as a viscoelastic. In addition, cell surface receptors have been identified that recognize the saccharides in hyaluronate, and interaction with specific proteins is responsible for the aggregate properties of connective tissue proteoglycans (see below). This diversity illustrates that a single polysaccharide may have both informational and physical roles in nature.
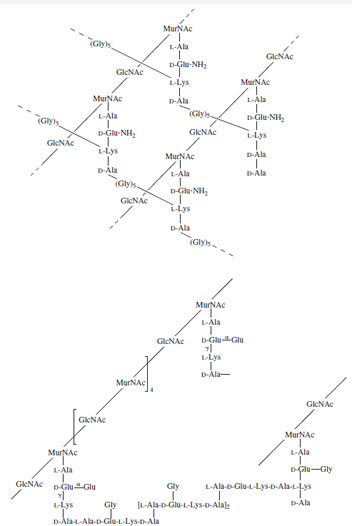 |
| Figure 23 Typical bacterial cell wall structures (peptidoglycans). |
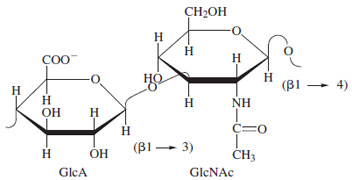 |
| Figure 24 Repeating unit of hyaluronate. This polysaccharide is distributed throughout connective tissue and is the only mammalian polysaccharide not covalently attached to protein. |
Support our developers




