Molecular Mimicry By Translational Factors
The mechanisms by which several of the translational factors act in protein synthesis have been suggested based on detailed structural analyses. Elongation factor Tu was the first factor for which an X-ray crystallographic structure was determined, and was also the first GTP-binding protein whose structure was elucidated. This protein is organized in three structural domains. Domain 1 is the GTPbinding domain (G-domain) consisting of a nucleotide binding fold now known to be typical of G-proteins. Domains 2 and 3 are β-barrels, held together by strong interdomain contacts such that the two domains act as a single structural unit. Crystal structures of the many functional forms of EF-Tu have been determined—EF-Tu:GDPNP (a nonhydrolyzable analog of GTP), EF-Tu:GDP, and complexes with EF-Ts or AA–tRNA. The orientation of Domain1withrespect toDomain2/3varies greatlyamong the functional forms of EF-Tu. The two parts of the protein rotate by 90◦ when GDP–GTP exchange is catalyzed by EF-Ts; this conformational change is due to two switch regions in the G-domain. By altering small secondary structure elements, these regions trigger long-range effects.The crystal structure of the ternary complex with Phe– tRNA (EF-Tu:GDPNP:Phe–tRNAPhe) demonstrated that the EF-Tu structure in the ternary complex is similar to that in the EF-Tu:GDPNP structure. Thus, binding of AA–tRNA does not alter the EF-Tu conformation. The ternary complex is quite elongated (Fig. 10), with the tRNA anticodon pointing away from EF-Tu, and close contacts are observed only between the factor and the Tstem, 3´-CCA–AA, and 5´-phosphate of the AA–tRNA. The Phe–tRNAPhe structure is also not significantly altered upon binding to EF-Tu.
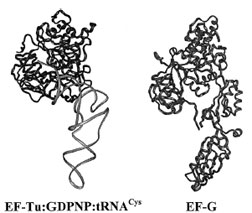 |
| Figure 10 Comparison of elongation factor structures. The crystal structures of the EF-Tu:GDPNP:tRNACys ternary complex (left) and EF-G (right) revealed that Domains 3, 4, and 5 of EF-G mimic the conformation of EF-Tu-bound tRNA. Several other translational factors have been determined or predicted to similarly mimic the tRNA structure. |
Upon comparison with the EF-Tu ternary complex structure, EF-G was seen to have a conformation similar to that of the ternary complex, with Domains 3, 4, and 5 of EF-G mimicking AA-tRNA.
The remarkable mimicry of tRNA by portions of EF-G suggested a mechanism by which the factor might be facilitating translocation. It is attractive to imagine that EF-G actively “chases” the peptidyl–tRNA into the ribosomal P-site because it mimics A-site-bound tRNA. Furthermore, the sequence of events dictates that, immediately after translocation, EF-G:GDP is released and another ternary complex (EF-Tu:GTP:AA–tRNA) enters the ribosomal A-site. Therefore the departure of EF-G:GDP leaves behind a binding site preformed for acceptance of the ternary complex.
Perhaps it should not be surprising that such structural mimicry is apparently used more than once in protein synthesis. Recent structure studies have determined that eRF1 (eukaryotic release Factor 1, which recognizes all stop codons) and RRF also contain domains that closely resemble tRNA. Although there are now several examples of factors that mimic the shape of tRNA, the factors themselves are structurally distinct from one another. This suggests a convergent appropriation of the tRNA structure, rather than a gene duplication event. In contrast, sequence comparisons of the G-protein factors EF-Tu, EF-G, IF2, and RF3 showed that the structural folds seen in Domains 1 and 2 of EF-Tu and EF-G are present in IF2 and RF3 and suggest that they will be present in the other factors as well.
Not only do several of the translational factors mimic the overall shape of tRNA, but also, even at the atomic level, a protein can copy the recognition events of the anticodon–codon interaction. The prokaryotic release factors RF1 and RF2 terminate polypeptide release at stop codons UAG/UAA and UGA/UAA, respectively. The basis for codon discrimination was localized to a tripeptide motif within the two release factors, with the first and third amino acids specifying the second and third base positions of the codon. For example, the first position (proline) of the “peptide anticodon” dictates that only A is recognized while serine is permissive to either A or G. Such relaxed recognition may be similar to wobble pairing in codon–anticodon interactions. This model for stop codon recognition was tested by generating protein variants that switched the codon specificity between release factors, recognized all stop codons, or were restricted to the UAA termination signal. The nucleotide feature being used for discrimination is likely the C2 amino group of G (where there is only a proton in A). The release factors are therefore making direct contact with the mRNA stop codons in a way that mimics the base-pairing of RNA–RNA interactions.
Support our developers




