Microscopic Structure of Bone
Microscopic Structure of Bone
Compact bone is composed of a calcified bone matrix arranged in concentric rings. The rings contain cavities (lacunae) filled with bone cells (osteocytes), which are interconnected by many minute passages (canaliculi). These passages serve to distribute nutrients throughout the bone. This entire organization of lacunae and canaliculi is arranged into an elongated cylinder called an osteon (also called haversian system) (Figure 31-7). Bone consists of bundles of osteons cemented together and interconnected with blood vessels and nerves. Because of blood vessels and nerves throughout bone, it is living tissue, although nonliving “ground substance” predominates. As a result of its living state, bone breaks can heal, and bone diseases can be as painful as any other tissue disease.
Following menopause, a woman loses 5% to 6% of her bone mass annually, often leading to the disease osteoporosis and increasing the risk of bone fractures. Dietary supplementation with calcium has been advocated to prevent such losses, but even large doses of calcium alone have little effect in slowing demineralization unless accompanied by therapy with the female sex hormone estrogen (because ovarian production of estrogen drops significantly after menopause). Among animals, only humans, especially females, are troubled with osteoporosis, perhaps a consequence of the long postreproductive life of the human species.
Bone growth is a complex restructuring process, involving both its destruction internally by boneresorbing cells (osteoclasts) and its deposition externally by bonebuilding cells (osteoblasts). Both processes occur simultaneously so that the marrow cavity inside grows larger by bone resorption while new bone is laid down outside by bone deposition. Bone growth responds to several hormones, in particular parathyroid hormone from the parathyroid gland, which stimulates bone resorption, and calcitonin from the thyroid gland, which inhibits bone resorption. These two hormones, together with a derivative of vitamin D, are responsible for maintaining a constant level of calcium in the blood. The effect of hormones on bone growth and resorption is described in more detail on.
Bone Like muscle, bone is subject to “use and disuse.” When we exercise our muscles, our bones respond by producing new bone tissue to give added strength. In fact, the bumps and processes to which muscles attach are produced by bone in response to the action of muscle forces. Conversely, when bones are not subject to stress, as in space flight, the body resorbs the mineral, and the bones become weak. Astronauts who spend many months in space must be carried from their capsules upon their return to earth.
Plan of the Vertebrate Skeleton
The vertebrate skeleton is composed of two main divisions: axial skeleton, which includes skull, vertebral column, sternum, and ribs, and appendicular skeleton, which includes the limbs (or fins or wings) and pectoral and pelvic girdles (Figures 31-8 and 31-9). Not surprisingly, the skeleton has undergone extensive remodeling in the course of vertebrate evolution. The move from water to land forced dramatic changes in body form. With increased cephalization, the further concentration of brain, sense organs, and food-gathering and respiratory apparatus in the head, the skull became the most intricate portion of the skeleton. Some early fishes had as many as 180 skull bones (a source of frustration to paleontologists) but through loss of some bones and fusion of others, skull bones became greatly reduced in number during evolution of the tetrapods. Amphibians and lizards have 50 to 95, and mammals, 35 or fewer. Humans have 29.
The vertebral column is the main stiffening axis of the postcranial skeleton. In fishes it serves much the same function as the notochord from which it is derived; that is, it provides points for muscle attachment and prevents telescoping of the body during muscle contraction. With evolution of amphibious and terrestrial tetrapods, the vertebrate body was no longer buoyed by the aquatic environment. The vertebral column became structurally adapted to withstand new regional stresses transmitted to the column by the two pairs of appendages. In amniote tetrapods (reptiles, birds, and mammals), the vertebrae are differentiated into cervical (neck), thoracic (chest), lumbar (back), sacral (pelvic), and caudal (tail) vertebrae. In birds and also in humans the caudal vertebrae are reduced in number and size, and the sacral vertebrae are fused. The number of vertebrae varies among the different vertebrates. Pythons seems to lead the list with more than 400. In humans (Figure 31-9) there are 33 in a young child, but in adults 5 are fused to form the sacrum and 4 to form the coccyx. Besides the sacrum and coccyx, humans have 7 cervical, 12 thoracic, and 5 lumbar vertebrae. The number of cervical vertebrae (7) is constant in nearly all mammals, whether the neck is short as in dolphins, or long as in giraffes.
The first two cervical vertebrae, atlas and axis, are modified to support the skull and permit pivotal movements. The atlas bears the globe of the head much as the mythological Atlas bore the earth on his shoulders. The axis, the second vertebra, permits the head to turn from side to side.
Ribs are long or short skeletal structures that articulate medially with vertebrae and extend into the body wall. Fishes have a pair of ribs for every vertebra (Figure 31-8); they serve as stiffening elements in the connective tissue septa that separate the muscle segments and thus improve the effectiveness of muscle contractions. Many fishes have both dorsal and ventral ribs, and some have numerous riblike intermuscular bones as well—all of which increase the difficulty and reduce the pleasure of eating certain kinds of fish. Other vertebrates have a reduced number of ribs, and some, such as the familiar leopard frog, have no ribs at all. In mammals the ribs together form the thoracic basket, which supports the chest wall and prevents collapse of the lungs. Mammals such as sloths have 24 pairs of ribs, whereas horses posses 18 pairs. Primates other than humans have 13 pairs of ribs; humans have 12 pairs, although approximately 1 person in 20 has a thirteenth pair.
Most vertebrates, fishes included, have paired appendages. All fishes except agnathans have thin pectoral and pelvic fins that are supported by the pectoral and pelvic girdles, respectively (Figure 31-8). Tetrapods (except caecilians, snakes, and limbless lizards) have two pairs of pentadactyl (fivetoed) limbs, also supported by girdles. The pentadactyl limb is similar in all tetrapods, alive and extinct; even when highly modified for various modes of life, the elements are rather easily homologized (the evolution of the pentadactyl limb is illustrated in Figure 27-1,).
Modifications of the basic pentadactyl limb for life in different environments involve distal elements much more frequently than proximal, and it is far more common for bones to be lost or fused than for new ones to be added. Horses and their relatives evolved a foot structure for fleetness by elongation of the third toe. In effect, a horse stands on its third fingernail (hoof), much like a ballet dancer standing on the tips of the toes. The bird wing is a good example of distal modification. The bird embryo bears 13 distinct wrist and hand bones (carpals and metacarpals), which are reduced to three digits in the adult. Most finger bones (phalanges) are lost, leaving four bones in three digits (see ). The proximal bones (humerus, radius, and ulna), however, are only slightly modified in the bird wing.
In nearly all tetrapods the pelvic girdle is firmly attached to the axial skeleton, since the greatest locomotory forces transmitted to the body come from the hindlimbs. The pectoral girdle, however, is much more loosely attached to the axial skeleton, providing the forelimbs with greater freedom for manipulative movements.
Effect of Body Size on Bone Stress
As Galileo realized in 1638, the ability of animals’ limbs to support a load decreases as animals increase in size (section opening essay,). Imagine two animals, one twice as long as the other, that are proportionally identical. That is, the larger animal is twice as long, twice as wide, and twice as tall as the smaller. The volume (and the weight) of the larger animal will be eight times the volume of the smaller (2 × 2 × 2 × 8). However, the strength of the larger animal’s legs will be only four times the strength of the smaller, because bone, tendon, and muscle strength are proportional to cross-sectional area. So, as Galileo noted, eight times the weight would have to be carried by only four times the strength. Because the maximum strength of mammalian bone is rather uniform per unit of cross-sectional area, how can animals become larger without placing unbearable stresses on long limb bones? One obvious solution is to make bones stouter and therefore stronger. However, throughout much of their size range, bone shape in different sized mammals does not change much. Instead, mammals have adapted limb posture so that stresses are shifted to align with the long axis of the bones, rather than transversely. Small animals the size of a chipmunk run in a crouched limb posture, whereas a large mammal such as a horse, has adopted an upright posture (Figure 31-10). Bones and muscles are capable of carrying far more weight when aligned more closely with the ground reaction force, as they are in a horse’s leg. In this way, peak bone stresses during strenuous activity are no greater for a galloping horse than for a running chipmunk or dog.
For animals larger than horses, further mechanical advantage by changing limb posture is not possible because the limbs are fully upright. Instead, the long bones of an elephant weighing 2.5 metric tons, and those of the enormous dinosaur Apatosaurus, weighing an estimated 34 metric tons, are (were) extremely thick and robust (Figure 31-10), providing the safety factor these massive animals require( d). However, top running speeds of the largest terrestrial mammals decline with increasing size. Nevertheless, recent calculations of bone stresses in dinosaurs suggest that even the largest were capable of considerable agility (Alexander, 1991).
Compact bone is composed of a calcified bone matrix arranged in concentric rings. The rings contain cavities (lacunae) filled with bone cells (osteocytes), which are interconnected by many minute passages (canaliculi). These passages serve to distribute nutrients throughout the bone. This entire organization of lacunae and canaliculi is arranged into an elongated cylinder called an osteon (also called haversian system) (Figure 31-7). Bone consists of bundles of osteons cemented together and interconnected with blood vessels and nerves. Because of blood vessels and nerves throughout bone, it is living tissue, although nonliving “ground substance” predominates. As a result of its living state, bone breaks can heal, and bone diseases can be as painful as any other tissue disease.
Following menopause, a woman loses 5% to 6% of her bone mass annually, often leading to the disease osteoporosis and increasing the risk of bone fractures. Dietary supplementation with calcium has been advocated to prevent such losses, but even large doses of calcium alone have little effect in slowing demineralization unless accompanied by therapy with the female sex hormone estrogen (because ovarian production of estrogen drops significantly after menopause). Among animals, only humans, especially females, are troubled with osteoporosis, perhaps a consequence of the long postreproductive life of the human species.
Bone growth is a complex restructuring process, involving both its destruction internally by boneresorbing cells (osteoclasts) and its deposition externally by bonebuilding cells (osteoblasts). Both processes occur simultaneously so that the marrow cavity inside grows larger by bone resorption while new bone is laid down outside by bone deposition. Bone growth responds to several hormones, in particular parathyroid hormone from the parathyroid gland, which stimulates bone resorption, and calcitonin from the thyroid gland, which inhibits bone resorption. These two hormones, together with a derivative of vitamin D, are responsible for maintaining a constant level of calcium in the blood. The effect of hormones on bone growth and resorption is described in more detail on.
Bone Like muscle, bone is subject to “use and disuse.” When we exercise our muscles, our bones respond by producing new bone tissue to give added strength. In fact, the bumps and processes to which muscles attach are produced by bone in response to the action of muscle forces. Conversely, when bones are not subject to stress, as in space flight, the body resorbs the mineral, and the bones become weak. Astronauts who spend many months in space must be carried from their capsules upon their return to earth.
Plan of the Vertebrate Skeleton
The vertebrate skeleton is composed of two main divisions: axial skeleton, which includes skull, vertebral column, sternum, and ribs, and appendicular skeleton, which includes the limbs (or fins or wings) and pectoral and pelvic girdles (Figures 31-8 and 31-9). Not surprisingly, the skeleton has undergone extensive remodeling in the course of vertebrate evolution. The move from water to land forced dramatic changes in body form. With increased cephalization, the further concentration of brain, sense organs, and food-gathering and respiratory apparatus in the head, the skull became the most intricate portion of the skeleton. Some early fishes had as many as 180 skull bones (a source of frustration to paleontologists) but through loss of some bones and fusion of others, skull bones became greatly reduced in number during evolution of the tetrapods. Amphibians and lizards have 50 to 95, and mammals, 35 or fewer. Humans have 29.
The vertebral column is the main stiffening axis of the postcranial skeleton. In fishes it serves much the same function as the notochord from which it is derived; that is, it provides points for muscle attachment and prevents telescoping of the body during muscle contraction. With evolution of amphibious and terrestrial tetrapods, the vertebrate body was no longer buoyed by the aquatic environment. The vertebral column became structurally adapted to withstand new regional stresses transmitted to the column by the two pairs of appendages. In amniote tetrapods (reptiles, birds, and mammals), the vertebrae are differentiated into cervical (neck), thoracic (chest), lumbar (back), sacral (pelvic), and caudal (tail) vertebrae. In birds and also in humans the caudal vertebrae are reduced in number and size, and the sacral vertebrae are fused. The number of vertebrae varies among the different vertebrates. Pythons seems to lead the list with more than 400. In humans (Figure 31-9) there are 33 in a young child, but in adults 5 are fused to form the sacrum and 4 to form the coccyx. Besides the sacrum and coccyx, humans have 7 cervical, 12 thoracic, and 5 lumbar vertebrae. The number of cervical vertebrae (7) is constant in nearly all mammals, whether the neck is short as in dolphins, or long as in giraffes.
5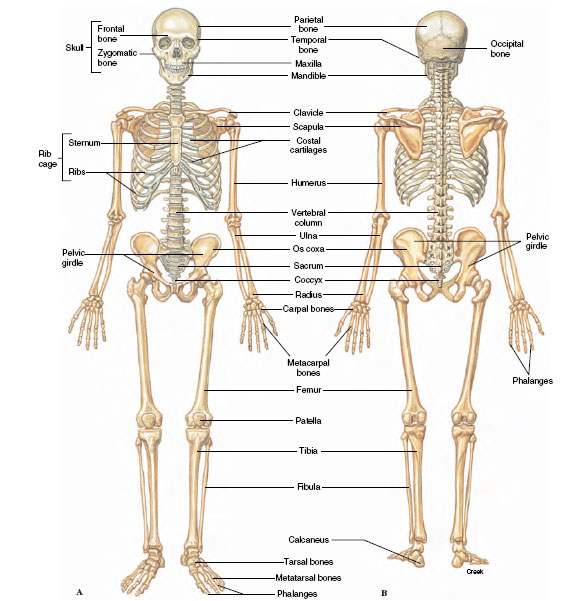 |
| Figure 31-9 Human skeleton. A, Ventral view. B, Dorsal view. In comparison with other mammals, the human skeleton is a patchwork of primitive and specialized parts. Erect posture, brought about by specialized changes in legs and pelvis, enabled the primitive arrangement of arms and hands (arboreal adaptation of human ancestors) to be used for manipulation of tools. Development of the skull and brain followed as a consequence of the premium natural selection put on dexterity and ability to appraise the environment. |
The first two cervical vertebrae, atlas and axis, are modified to support the skull and permit pivotal movements. The atlas bears the globe of the head much as the mythological Atlas bore the earth on his shoulders. The axis, the second vertebra, permits the head to turn from side to side.
Ribs are long or short skeletal structures that articulate medially with vertebrae and extend into the body wall. Fishes have a pair of ribs for every vertebra (Figure 31-8); they serve as stiffening elements in the connective tissue septa that separate the muscle segments and thus improve the effectiveness of muscle contractions. Many fishes have both dorsal and ventral ribs, and some have numerous riblike intermuscular bones as well—all of which increase the difficulty and reduce the pleasure of eating certain kinds of fish. Other vertebrates have a reduced number of ribs, and some, such as the familiar leopard frog, have no ribs at all. In mammals the ribs together form the thoracic basket, which supports the chest wall and prevents collapse of the lungs. Mammals such as sloths have 24 pairs of ribs, whereas horses posses 18 pairs. Primates other than humans have 13 pairs of ribs; humans have 12 pairs, although approximately 1 person in 20 has a thirteenth pair.
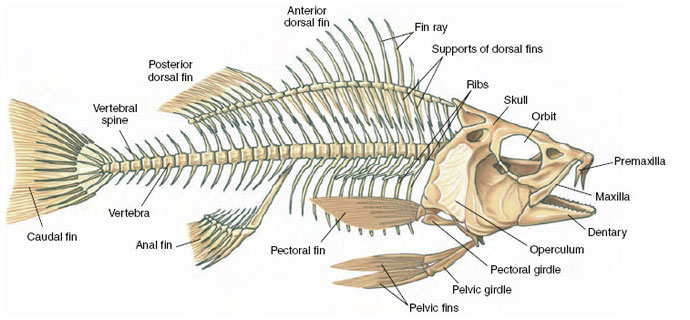 |
| Figure 31-8 Skeleton of a perch |
Most vertebrates, fishes included, have paired appendages. All fishes except agnathans have thin pectoral and pelvic fins that are supported by the pectoral and pelvic girdles, respectively (Figure 31-8). Tetrapods (except caecilians, snakes, and limbless lizards) have two pairs of pentadactyl (fivetoed) limbs, also supported by girdles. The pentadactyl limb is similar in all tetrapods, alive and extinct; even when highly modified for various modes of life, the elements are rather easily homologized (the evolution of the pentadactyl limb is illustrated in Figure 27-1,).
Modifications of the basic pentadactyl limb for life in different environments involve distal elements much more frequently than proximal, and it is far more common for bones to be lost or fused than for new ones to be added. Horses and their relatives evolved a foot structure for fleetness by elongation of the third toe. In effect, a horse stands on its third fingernail (hoof), much like a ballet dancer standing on the tips of the toes. The bird wing is a good example of distal modification. The bird embryo bears 13 distinct wrist and hand bones (carpals and metacarpals), which are reduced to three digits in the adult. Most finger bones (phalanges) are lost, leaving four bones in three digits (see ). The proximal bones (humerus, radius, and ulna), however, are only slightly modified in the bird wing.
In nearly all tetrapods the pelvic girdle is firmly attached to the axial skeleton, since the greatest locomotory forces transmitted to the body come from the hindlimbs. The pectoral girdle, however, is much more loosely attached to the axial skeleton, providing the forelimbs with greater freedom for manipulative movements.
Effect of Body Size on Bone Stress
As Galileo realized in 1638, the ability of animals’ limbs to support a load decreases as animals increase in size (section opening essay,). Imagine two animals, one twice as long as the other, that are proportionally identical. That is, the larger animal is twice as long, twice as wide, and twice as tall as the smaller. The volume (and the weight) of the larger animal will be eight times the volume of the smaller (2 × 2 × 2 × 8). However, the strength of the larger animal’s legs will be only four times the strength of the smaller, because bone, tendon, and muscle strength are proportional to cross-sectional area. So, as Galileo noted, eight times the weight would have to be carried by only four times the strength. Because the maximum strength of mammalian bone is rather uniform per unit of cross-sectional area, how can animals become larger without placing unbearable stresses on long limb bones? One obvious solution is to make bones stouter and therefore stronger. However, throughout much of their size range, bone shape in different sized mammals does not change much. Instead, mammals have adapted limb posture so that stresses are shifted to align with the long axis of the bones, rather than transversely. Small animals the size of a chipmunk run in a crouched limb posture, whereas a large mammal such as a horse, has adopted an upright posture (Figure 31-10). Bones and muscles are capable of carrying far more weight when aligned more closely with the ground reaction force, as they are in a horse’s leg. In this way, peak bone stresses during strenuous activity are no greater for a galloping horse than for a running chipmunk or dog.
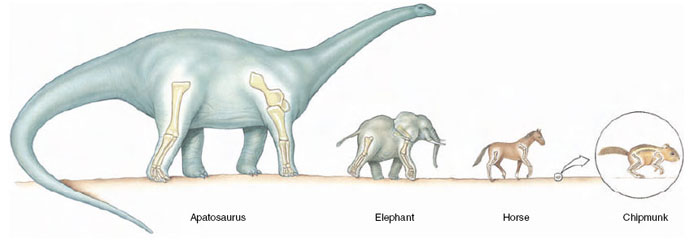 |
| Figure 31-10 Comparison of postures in small and large mammals, showing the effect of scale. Because of its more upright posture, bone stresses in the horse are similar to those in the chipmunk. In mammals larger than horses (above about 300 kg), greatly increased stresses require that bones become exceedingly robust and that the animal lose agility. |
For animals larger than horses, further mechanical advantage by changing limb posture is not possible because the limbs are fully upright. Instead, the long bones of an elephant weighing 2.5 metric tons, and those of the enormous dinosaur Apatosaurus, weighing an estimated 34 metric tons, are (were) extremely thick and robust (Figure 31-10), providing the safety factor these massive animals require( d). However, top running speeds of the largest terrestrial mammals decline with increasing size. Nevertheless, recent calculations of bone stresses in dinosaurs suggest that even the largest were capable of considerable agility (Alexander, 1991).
Support our developers




