Vertebrate Integument and Derivatives
Vertebrate Integument and Derivatives
The basic plan of the vertebrate integument, as exemplified by frog and human skin (Figure 31-1B and C), includes a thin, outer stratified epithelial layer, the epidermis, derived from ectoderm and an inner, thicker layer, the dermis, or true skin, which is of mesodermal origin.
Although the epidermis is thin and appears simple in structure, it gives rise to most derivatives of the integument, such as hair, feathers, claws, and hooves. The dermis contains blood vessels, collagenous fibers, nerves, pigment cells, fat cells, and connective tissue cells called fibroblasts. These elements support, cushion, and nourish the epidermis, which is devoid of blood vessels.
The epidermis is a stratified squamous epithelium consisting usually of several layers of cells. The basal part is made up of cells that undergo frequent mitosis to renew layers that lie above. As outer layers of cells are displaced upward by new generations of cells beneath, an exceedingly tough, fibrous protein called keratin accumulates in the interior of the cells. Gradually, keratin replaces all metabolically active cytoplasm. The cell dies and is eventually shed, lifeless and scalelike. Such is the origin of dandruff as well as a significant fraction of household dust. This process is called keratinization, and the cell, thus transformed, is said to be cornified. Cornified cells, highly resistant to abrasion and water diffusion, comprise the outermost stratum corneum. This epidermal layer becomes especially thick in areas exposed to persistent pressure or wear such as calluses, foot pads of mammals, and the scales of reptiles and birds. The dermis, as already mentioned, mainly serves a supportive role for the epidermis. Nevertheless, true bony structures, where they occur in the integument, are always dermal derivatives. Heavy bony plates were common in ostracoderms and placoderms of the Paleozoic era (Figure 25-17,) and persist is some living fishes, such as sturgeons (Figure 26-19,). Scales of contemporary fishes are bony dermal structures that have evolved from the bony armor of the Paleozoic fishes but are much smaller and more flexible. Fish scales are thin bony slivers covered with a mucus-secreting epidermis (Figure 31-2). Most amphibians lack dermal bones in their skin, whereas in reptiles dermal bones provide the armor of crocodilians, the beaded skin appearance of many lizards, and also contribute to the shell of turtles. Dermal bone also gives rise to antlers, as well as the bony core of horns.
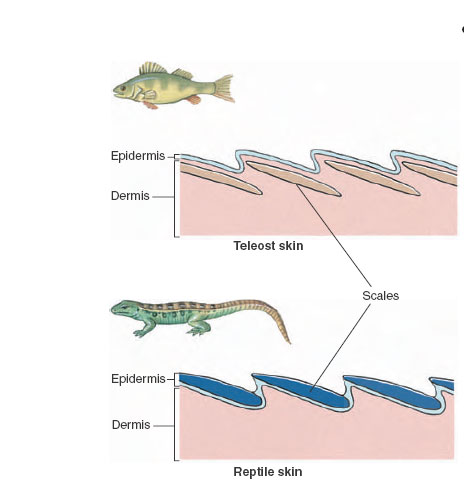 |
| Figure 31-2 Integument of bony fishes and lizards. Bony (teleost) fishes have bony scales from dermis, and lizards have horny scales from epidermis. Thus they are not homologous structures. Dermal scales of fishes are retained throughout life. Since a new growth ring is added to each scale each year, fishery biologists use scales to tell the age of fishes. Epidermal scales of reptiles are shed periodically. |
Lizards, snakes, turtles, and crocodilians were among the first to exploit the adaptive possibilities of the remarkably tough protein keratin.The reptilian epidermal scale that develops from keratin is a much lighter and more flexible structure than the bony, dermal scale of fishes, yet it provides excellent protection from abrasion and desiccation. Scales may be overlapping structures, as in snakes and some lizards, or develop into plates, as in turtles and crocodilians. In birds, keratin found new uses. Feathers, beaks, and claws, as well as scales, are all epidermal structures composed of dense keratin. Mammals continued to capitalize on keratin’s virtues by turning it into hair, hooves, claws, and nails.As a result of its keratin content, hair is by far the strongest material in the body. It has a tensile strength comparable to that of rolled aluminum and is nearly twice as strong, weight for weight, as the strongest bone.
 |
| Figure 31-3
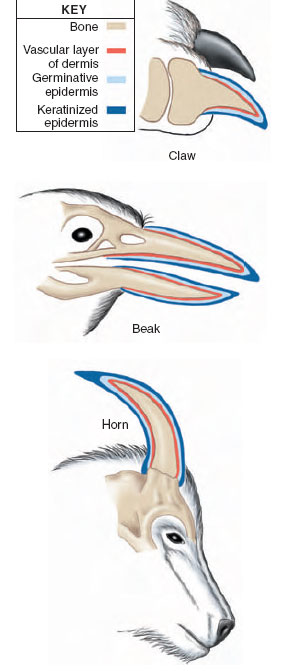
Similarity of structure of integumentary derivatives. Claws, beaks, and horns are all built of similar combinations of epidermal (keratinized) and dermal components. A central bony core is covered by a vascularized nutritive layer of the dermis. An outer epithelial layer has a basal germinative component which proliferates to allow these structures to grow continually. The thickened surface epithelium is keratinized or cornified. Note that the relative thickness of each component is not drawn to scale. |
Structures such as claws, beaks, nails, and horns are made up of combinations of epidermal (keratinized) and dermal components. Their basic structure is the same, with a central bony core covered by a vascularized nutritive layer of the dermis, and an outer epithelial layer. This epithelial layer has a germinative component responsible for the continual growth of horns, hooves, claws, and beaks. The outer epithelial layer is keratinized. Overgrowth of these structures is prevented by constant wear and abrasion (Figure 31-3).
Animal Coloration
The colors of animals may be vivid and dramatic when serving as important recognition marks or as warning coloration, or they may be subdued or cryptic when used for camouflage. Integumentary color is usually produced by pigments, but in many insects and in some vertebrates, especially birds, certain colors are produced by the physical structure of the surface tissue, which reflects certain light wavelengths and eliminates others. Colors produced this way are called structural color, and they are responsible for the most beautifully iridescent and metallic hues to be found in the animal kingdom. Many butterflies and beetles and a few fishes thus share with birds the distinction of being the earth’s most resplendent animals. Certain structural colors of feathers are caused by minute, air-filled spaces or pores that reflect white light (white feathers) or some portion of the spectrum (for example, Tyndall blue coloration produced by scattering of light [see note, ]). Iridescent colors that change hue as the animal’s angle shifts with respect to the observer are produced when light is reflected from several layers of thin, transparent film. By phase interference, light waves reinforce, weaken, or eliminate each other to produce some of the purest and most brilliant colors we know.
More common than structural colors in animals are pigments (biochromes), an extremely varied group of large molecules that reflect light rays. In crustaceans and ectothermic vertebrates these pigments are contained in large cells with branching processes, called chromatophores (Figure 31-4A). The pigment may concentrate in the center of the cell in an aggregate too small to be visible, or it may disperse throughout the cell and its processes, providing maximum display. The chromatophores of cephalopod molluscs are entirely different (Figure 31-4B). Each is a small sac-like cell filled with pigment granules and surrounded by muscle cells that, when contracted, stretch the whole cell out into a pigmented sheet. When the muscles relax, the elastic chromatophore quickly shrinks to a small sphere. With such pigment cells the squids and octopuses can alter their color more rapidly than any other animal.
The most widespread of animal pigments are the melanins, a group of black or brown polymers that are responsible for the various earthcolored shades that most animals wear. Yellow and red colors are often caused by carotenoid pigments, which are frequently contained within special pigment cells called xanthophores. Most vertebrates are incapable of synthesizing their own carotenoid pigments but must obtain them directly or indirectly from plants. Two entirely different classes of pigments called ommochromes and pteridines are usually responsible for the yellow pigments of molluscs and arthropods. Green colors are rare; when they occur, they are usually produced by yellow pigment overlying blue structural color. Iridophores, a third type of chromatophore, contain crystals of guanine or some other purine, rather than pigment. They produce a silvery or metallic effect by reflecting light.
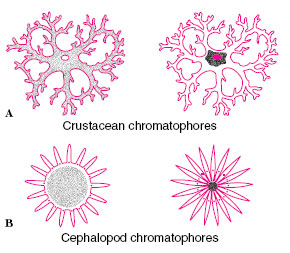 |
| Figure 31-4 Chromatophores. A,The crustacean chromatophore showingthe pigment dispersed (left) and concentrated (right). Vertebrate chromatophores are similar. B, The cephalopod chromatophore is an elastic capsule surrounded by musclefibers that, when contracted (left), stretch out the capsule to expose the pigment. |
By vertebrate standards, mammals are a somber-colored group. Most mammals are more or less color blind, a deficiency that is doubtless connected with the lack of bright colors in the group. Exceptions are the brilliantly colored skin patches of some baboons and mandrills. Significantly, primates have color vision and thus can appreciate such eye-catching ornaments. The muted colors of mammals are caused by melanin, which is deposited in growing hair by dermal melanophores.
Injurious Effects of Sunlight
The familiar vulnerability of the human skin to sunburn reminds us of the potentially damaging effects of ultraviolet radiation on protoplasm. Many animals, such as protozoa and flatworms, if exposed to the sun in shallow water are damaged or killed by ultraviolet radiation. Most land animals are protected from such damage by the screening action of special body coverings, for example, the cuticle of arthropods, the scales of reptiles, and the feathers and fur of birds and mammals. Humans, however, are “naked apes” that lack the furry protection of most other mammals. We must depend on thickening of the epidermis (stratum corneum) and on epidermal pigmentation for protection. Most ultraviolet radiation is absorbed in the epidermis, but about 10% penetrates the dermis. Damaged cells in both the epidermis and dermis release histamine and other vasodilator substances that cause blood vessel enlargement in the dermis and the characteristic red coloration of sunburn. Light skins suntan through the formation of the pigment melanin in the deeper epidermis and by “pigment darkening,” that is, the photooxidative blackening of bleached pigment already present in the epidermis. Unfortunately, tanning does not bestow perfect protection. Sunlight still ages the skin prematurely, and tanning itself causes the skin to become dry and leathery. Sunlight also is responsible for approximately 1 million new cases of skin cancer annually in the United States alone, making skin cancer the most common of malignancies among Caucasians. There is now strong evidence that genetic mutations caused by high doses of sunlight received during the pre-adult years are responsible for skin cancers that appear after middle age.
Support our developers




