Rigid Skeletons
Rigid Skeletons
Rigid skeletons differ from hydrostatic skeletons in one fundamental way: rigid skeletons consist of rigid elements, usually jointed, to which muscles can attach. Muscles can only contract; to be lengthened they must be extended by the pull of an antagonistic set of muscles. Rigid skeletons provide the anchor points required by opposing sets of muscles, such as flexors and extensors.
Antagonistic muscles are functional opposites that oppose each other’s action. For example, the biceps brachii on one side of the upper arm is opposed in its action by the triceps brachii on the opposite side of the arm. By contracting against each other, they balance and smooth rapid movements.
There are two principal types of rigid skeletons: exoskeleton, typical of molluscs, arthropods and many other invertebrates; and endoskeleton, characteristic of echinoderms and vertebrates. The invertebrate exoskeleton may be mainly protective, but it may also perform a vital role in locomotion. An exoskeleton may take the form of a shell, a spicule, or a calcareous, proteinaceous, or chitinous plate. It may be rigid, as in molluscs, or jointed and movable, as in arthropods. Unlike an endoskeleton, which grows with the animal, an exoskeleton is often a limiting coat of armor that must be periodically molted to make way for an enlarged replacement (molting in crustaceans is described on). Some invertebrate exoskeletons, such as the shells of snails and bivalves, grow with the animal.
The arthropod-type exoskeleton is perhaps a better arrangement for small animals than a vertebrate-type endoskeleton because a hollow cylindrical tube can support much more weight without collapsing than can a solid cylindrical rod of the same material and weight. Arthropods can thus enjoy both protection and structural support from their exoskeleton. But for larger animals the hollow cylinder would be completely impractical. If made thick enough to support the body weight, it would be too heavy to lift; but if kept thin and light, it would be extremely sensitive to buckling or shattering on impact. Finally, can you imagine the sad plight of an animal the size of an elephant when it shed its exoskeleton to molt?
The vertebrate endoskeleton is formed inside the body and is composed of bone and cartilage, which are forms of dense connective tissue. Bone not only supports and protects but is also the major body reservoir for calcium and phosphorus. In amniote vertebrates red blood cells and certain white blood cells are formed in the bone marrow.
Notochord and Cartilage
The notochord (see Figure 25-1, ) is a semirigid supportive axial rod of the protochordates and all vertebrate larvae and embryos. It is composed of large, vacuolated cells and is surrounded by layers of elastic and fibrous sheaths. It is a stiffening device, preserving body shape during locomotion. Except in the jawless vertebrates (lampreys and hagfishes), the notochord is surrounded or replaced by the backbone during embryonic development.
Cartilage is a major skeletal element of some vertebrates. The jawless fishes (for example, lampreys) and the elasmobranchs (sharks, skates, and rays) have purely cartilaginous skeletons, which oddly enough is a derived feature, since their Paleozoic ancestors had bony skeletons. Other vertebrates as adults have principally bony skeletons with some cartilage interspersed. Cartilage is a soft, pliable, characteristically deep-lying tissue. Unlike most connective tissues, which are quite variable in form, cartilage is basically the same wherever it is found. The basic form, hyaline cartilage, has a clear, glassy appearance (see Figure 9-6,). It is composed of cartilage cells (chondrocytes) surrounded by firm complex protein gel interlaced with a meshwork of collagenous fibers. Blood vessels are virtually absent—the reason that sports injuries involving cartilage heal poorly. In addition to forming the cartilaginous skeleton of some vertebrates and that of all vertebrate embryos, hyaline cartilage makes up the articulating surfaces of many bone joints of most adult vertebrates and the supporting tracheal, laryngeal, and bronchial rings.
Cartilage similar to hyaline cartilage occurs in some invertebrates, for example the radula of gastropod molluscs and lophophore of brachiopods. The cartilage of cephalopod molluscs is of a special type with long, branching processes that resemble the cells of vertebrate bone.
Bone
Bone is a living tissue that differs from other connective and supportive tissues by having significant deposits of inorganic calcium salts laid down in an extracellular matrix. Its structural organization is such that bone has nearly the tensile strength of cast iron, yet is only one-third as heavy.
Bone is never formed in vacant space but is always laid down by replacement in areas occupied by some form of connective tissue. Most bone develops from cartilage and is called endochondral (“withincartilage”) or replacement bone. Embryonic cartilage is gradually eroded leaving it extensively honeycombed; bone-forming cells then invade these areas and begin depositing calcium salts around strandlike remnants of the cartilage. A second type of bone is intramembranous bone, which develops directly from sheets of embryonic cells. Dermal bone, mentioned earlier, is a type of intramembranous bone. In tetrapod vertebrates intramembranous bone is restricted mainly to bones of the face, cranium and clavicle; the remainder of the skeleton is endochondral bone. Whatever the embryonic origin, once fully formed, endochondral and intramembranous bone look the same.
Fully formed bone, however, may vary in density. Cancellous (or spongy) bone consists of an open, interlacing framework of bony tissue, oriented to give maximum strength under the normal stresses and strains that the bone receives. All bone develops first as cancellous bone, but some bones, through further deposition of bone salts, become compact. Compact bone is dense, appearing solid to the unaided eye. Both cancellous and compact bone are found in the typical long bones of tetrapods (Figure 31-7).
Rigid skeletons differ from hydrostatic skeletons in one fundamental way: rigid skeletons consist of rigid elements, usually jointed, to which muscles can attach. Muscles can only contract; to be lengthened they must be extended by the pull of an antagonistic set of muscles. Rigid skeletons provide the anchor points required by opposing sets of muscles, such as flexors and extensors.
Antagonistic muscles are functional opposites that oppose each other’s action. For example, the biceps brachii on one side of the upper arm is opposed in its action by the triceps brachii on the opposite side of the arm. By contracting against each other, they balance and smooth rapid movements.
There are two principal types of rigid skeletons: exoskeleton, typical of molluscs, arthropods and many other invertebrates; and endoskeleton, characteristic of echinoderms and vertebrates. The invertebrate exoskeleton may be mainly protective, but it may also perform a vital role in locomotion. An exoskeleton may take the form of a shell, a spicule, or a calcareous, proteinaceous, or chitinous plate. It may be rigid, as in molluscs, or jointed and movable, as in arthropods. Unlike an endoskeleton, which grows with the animal, an exoskeleton is often a limiting coat of armor that must be periodically molted to make way for an enlarged replacement (molting in crustaceans is described on). Some invertebrate exoskeletons, such as the shells of snails and bivalves, grow with the animal.
1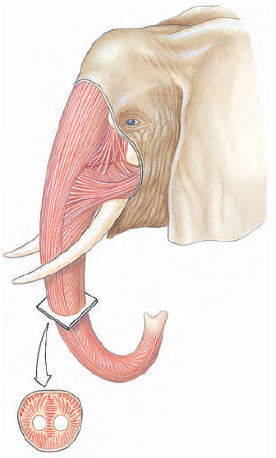 |
| Figure 31-6
Muscular trunk of an elephant, an example of a muscular hydrostat. |
The arthropod-type exoskeleton is perhaps a better arrangement for small animals than a vertebrate-type endoskeleton because a hollow cylindrical tube can support much more weight without collapsing than can a solid cylindrical rod of the same material and weight. Arthropods can thus enjoy both protection and structural support from their exoskeleton. But for larger animals the hollow cylinder would be completely impractical. If made thick enough to support the body weight, it would be too heavy to lift; but if kept thin and light, it would be extremely sensitive to buckling or shattering on impact. Finally, can you imagine the sad plight of an animal the size of an elephant when it shed its exoskeleton to molt?
The vertebrate endoskeleton is formed inside the body and is composed of bone and cartilage, which are forms of dense connective tissue. Bone not only supports and protects but is also the major body reservoir for calcium and phosphorus. In amniote vertebrates red blood cells and certain white blood cells are formed in the bone marrow.
Notochord and Cartilage
The notochord (see Figure 25-1, ) is a semirigid supportive axial rod of the protochordates and all vertebrate larvae and embryos. It is composed of large, vacuolated cells and is surrounded by layers of elastic and fibrous sheaths. It is a stiffening device, preserving body shape during locomotion. Except in the jawless vertebrates (lampreys and hagfishes), the notochord is surrounded or replaced by the backbone during embryonic development.
Cartilage is a major skeletal element of some vertebrates. The jawless fishes (for example, lampreys) and the elasmobranchs (sharks, skates, and rays) have purely cartilaginous skeletons, which oddly enough is a derived feature, since their Paleozoic ancestors had bony skeletons. Other vertebrates as adults have principally bony skeletons with some cartilage interspersed. Cartilage is a soft, pliable, characteristically deep-lying tissue. Unlike most connective tissues, which are quite variable in form, cartilage is basically the same wherever it is found. The basic form, hyaline cartilage, has a clear, glassy appearance (see Figure 9-6,). It is composed of cartilage cells (chondrocytes) surrounded by firm complex protein gel interlaced with a meshwork of collagenous fibers. Blood vessels are virtually absent—the reason that sports injuries involving cartilage heal poorly. In addition to forming the cartilaginous skeleton of some vertebrates and that of all vertebrate embryos, hyaline cartilage makes up the articulating surfaces of many bone joints of most adult vertebrates and the supporting tracheal, laryngeal, and bronchial rings.
Cartilage similar to hyaline cartilage occurs in some invertebrates, for example the radula of gastropod molluscs and lophophore of brachiopods. The cartilage of cephalopod molluscs is of a special type with long, branching processes that resemble the cells of vertebrate bone.
Bone
Bone is a living tissue that differs from other connective and supportive tissues by having significant deposits of inorganic calcium salts laid down in an extracellular matrix. Its structural organization is such that bone has nearly the tensile strength of cast iron, yet is only one-third as heavy.
Bone is never formed in vacant space but is always laid down by replacement in areas occupied by some form of connective tissue. Most bone develops from cartilage and is called endochondral (“withincartilage”) or replacement bone. Embryonic cartilage is gradually eroded leaving it extensively honeycombed; bone-forming cells then invade these areas and begin depositing calcium salts around strandlike remnants of the cartilage. A second type of bone is intramembranous bone, which develops directly from sheets of embryonic cells. Dermal bone, mentioned earlier, is a type of intramembranous bone. In tetrapod vertebrates intramembranous bone is restricted mainly to bones of the face, cranium and clavicle; the remainder of the skeleton is endochondral bone. Whatever the embryonic origin, once fully formed, endochondral and intramembranous bone look the same.
Fully formed bone, however, may vary in density. Cancellous (or spongy) bone consists of an open, interlacing framework of bony tissue, oriented to give maximum strength under the normal stresses and strains that the bone receives. All bone develops first as cancellous bone, but some bones, through further deposition of bone salts, become compact. Compact bone is dense, appearing solid to the unaided eye. Both cancellous and compact bone are found in the typical long bones of tetrapods (Figure 31-7).
5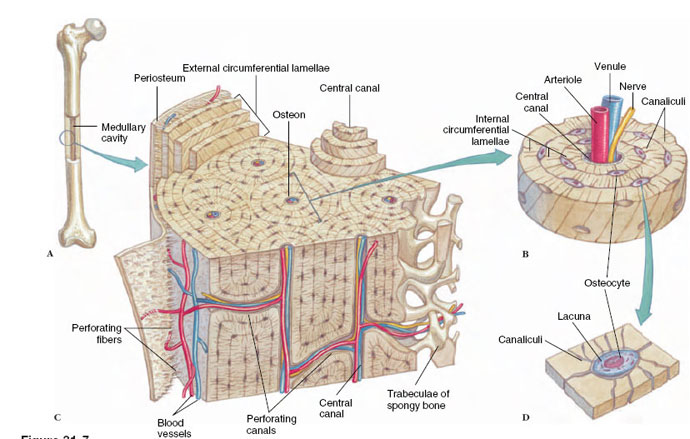 |
| Figure 31-7 Structure of compact bone. A, Adult long bone with a cut into the medullary cavity. B, Enlarged section showing osteons, the basic histological unit of bone. C, Enlarged view of an osteon showing the concentric lamellae and the osteocytes (bone cells) arranged within lacunae. D, An osteocyte within a lacuna. Bone cells receive nutrients from the circulatory system via tiny canaliculi that interlace the calcified matrix. Bone cells are known as osteoblasts when they are building bone, but, in mature bone shown here, they become resting osteocytes. Bone is covered with compact connective tissue called periosteum. |
Support our developers




